Атф синтезируется не в митохондриях кишечной палочки

Американские биологи заставили кишечную палочку поселиться внутри клеток дрожжей и выполнять функции сломанных митохондрий. Результаты эксперимента, который имитирует ранние этапы эволюции митохондрий, описаны в статье, опубликованной в журнале Proceedings of the National Academy of Sciences.
Митохондрия – важнейшая органелла клеток человека, а заодно и практически всех других эукариотических организмов, главная энергетическая станция клетки. На ней углеводы окисляются под воздействием кислорода, а выделившаяся при этом энергия запасается впрок. Из цитоплазмы туда поступают «разряженные» клеточные «батарейки» – молекулы АДФ. Там они «заряжаются», превращаясь в молекулы АТФ, покидают митохондрию и используются дальше на нужды клетки.
Еще в начале XX века ученые заметили, что митохондрии (и хлоропласты растений) удивительно похожи на бактерий, а в 1970-х годах Линн Маргулис и ее последователи свели сведения об этом в теорию эндосимбиоза. Согласно ей, все митохондрии были раньше свободноживущими бактериями, способными очень эффективно перерабатывать углеводы при помощи кислорода, а потом попали внутрь предковой эукариотической клетки. По каким-то причинам они не были переварены, как это обычно происходит, а остались целы. Клетка-хозяин предоставила им укрытие, стала снабжать разными необходимыми веществами, а симбионты, в свою очередь, стали снабжать клетку энергией.
С того момента бывшая свободноживущая бактерия сильно видоизменилась и настолько приспособилась к жизни внутри клетки, что теперь считается ее частью. Например, в митохондриях осталась лишь короткая ДНК с самым базовым набором генов, тогда как большая их часть исчезла за ненадобностью или переехала жить в ядерный геном. Поскольку симбиоз случился довольно давно – больше полутора миллиардов лет назад, – разобраться в ранних этапах со-настройки предков эукариот и их митохондрий времена довольно сложно. В основном это делается при помощи реконструкции на основе современных митохондрий.
Ангад Мехта (Angad Mehta) из Института Скриппс и его коллеги из нескольких калифорнийских институтов подошли к решению этого вопроса с неожиданной стороны. Они предложили создать для исследований синтетическую модель свежего эндосимбиоза бактерий и эукариот, «подружив» два современных свободноживущих организма: дрожжи (Saccharomyces cerevisiae) и кишечную палочку (Escherichia coli). Для получения корректной и жизнеспособной модели оказалось необходимым правильно подготовить встречу потенциальных симбионтов.
Для начала клетки дрожжей «заинтересовали» в симбиозе. Поскольку у современных эукариот уже есть митохондрии, исследователи попытались их «отключить». Для эксперимента сперва взяли клетки дрожжей, у которых вообще не было митохондриальной ДНК. Поскольку в ней закодировано все самое нужное для работы митохондрий, то такие клетки жили очень плохо и только на специальной питательной среде, не требующей переработки углеводов митохондриями. Кроме этого, ученые взяли дрожжи, в которых митохондрии были покалечены чуть меньше – в них испортили всего один, но важный ген cox2.
В пару к дрожжам была подобран симбионт – такой, в присутствие которого которого эти поломки бы компенсировались. Кишечная палочка Esherichia coli – модельная бактерия в биологии – относительно близкий родственник предков митохондрий. Тем не менее, ее тоже пришлось адаптировать чтобы научить дорожить дружбой с дрожжами. Во-первых, ей сломали путь биосинтеза тиамина (он же витамин B1). Теперь она могла расти только в среде, в которой он есть, например с дрожжами. Во-вторых, её заставили делиться энергией с потенциальным хозяином. В качестве аккумуляторов энергии клетки используют АТФ, и в нормальных эукариотических клетках в мембрану митохондрии встроен транспортер, который позволяет АТФ проходить из митохондрии в цитоплазму, а молекулам АДФ («разряженный аккумулятор») обратно. У кишечной палочки такого транспортера не предусмотрено, и поэтому его пришлось вставить его из другой бактерии. Помимо того, кишечной палочке добавили ген зеленого флуоресцентного белка чтобы бактерию было легко различить с помощью микроскопии.
После того, как организмы были подготовлены ко встрече друг с другом, их посадили вместе, и дальше, чтобы отобрать варианты с удачным симбиозом, начали растить на среде, для переработки которой дрожжам бы понадобилась помощь кишечных палочек. Оказалось, что дрожжи без митохондриальной ДНК не выживают в таких условиях, а дрожжи с выключенным cox2 геном образуют очень маленькое число колоний. По идее, эти колонии могли выжить за счет бактерий, которые их окружали, и не формировать эндосимбиоз. Чтобы учесть такой вариант, исследователи пересадили эти колонии на среду, в которой не могут расти отдельно ни дрожжи, ни кишечные палочки (без тиамина и без альтернативных источников энергии). Оказалось, что в таких условиях не выжил никто.
Чтобы еще больше увеличить вероятность удачного симбиоза, исследователи решили повлиять на способность бактерий не перевариваться внутри дрожжей. У патогенных бактерий есть для этой цели специальные SNARE-подобные белки. Они помогают манипулировать везикулярным транспортом хозяина и избежать попадания в лизосому – «желудок клетки». Кишечную палочку, которая к этому моменту уже синтезировали флуоресцентный белок и светилась зеленым светом, не росла без тиамина и могла выпускать АТФ наружу клетки, исследователи наделили вдобавок SNARE-подобными генами из патогенной хламидии, и повторили эксперимент еще раз. На этот раз было обнаружено много химерных организмов из дрожжевой клетки с бактериями внутри, которые поддерживались на протяжении нескольких поколений. Чтобы полностью исключить возможность внешнего симбиоза, исследователи добавили в среду антибиотик, но и в этом случае колонии химер не исчезали даже после нескольких раундов пересаживания со среды на среду.
Авторы статьи считают, что подобная синтетическая модель эндосимбиоза позволит лучше разобраться в том, как происходила коэволюция митохондрии и эукариотической клетки, в том числе – как уменьшался бактериальный геном (свободноживущая бактерия обычно имеет несколько тысяч разных генов, тогда как в геноме митохондрии их не больше ста). Например, они попробовали убрать из него еще один важный ген и оказалось, что химеры с такими кишечными палочками тоже вполне жизнеспособны.
Вера Мухина
Источник
Каждое живое существо должно получать энергию из окружающей среды (например, в форме солнечного излучения или органических продуктов питания). Эта энергия требует для биосинтеза (анаболизма) огромного числа химических соединений и биополимеров в соответствии с определенной генетической программой. Сама энергия нужна для активной передачи молекул и ионов через мембраны, для движения и для передачи нервных импульсов. Наука, которая изучает поток и использование энергии в живых существах, называется «биоэнергетика» (био- + энергия).
Роль АТФ в энергетическом балансе
АТФ является основной молекулой энергии в живых системах. Он участвует в различных химических процессах, от химического биосинтеза до движения ресничек, сокращения мышц, активного транспорта молекул через клеточную мембрану или распространения электрического импульса через нервные волокна.
Производство и потребление энергии происходит через сеть ферментативных реакций (метаболизм). Центральным химическим соединением в метаболизме является аденозинтрифосфат (АТФ), который образуется в результате метаболических реакций (катаболизм) путем фосфорилирования аденозиндифосфата (АДФ) с образованием энергии около 30 кДж / моль (термодинамика). Большая часть АТФ производится в результате процессов в митохондриях (окислительного фосфорилирования). При использовании этой энергии в биологических процессах АТФ обычно гидролизуется до фосфата и АДФ (аденозинДИфосфата).
Вся биосинтетическая деятельность, как и многие другие клеточные действия, требует энергии. В основном для клеточных активностей источником энергии является именно АТФ. Молекула АТФ состоит из аденина, рибозы и трех фосфатных групп (ФГ). Последние с сильным отрицательным зарядом связаны двумя ковалентными высокоэнергетическими связями, которые при гидролизе выделяют относительно много энергии. Это демонстрирует важное свойство АТФ.
Поэтому без преувеличения можно сказать, что наиболее важным энергетическим соединением в клетке является трифосфат аденозина (АТФ), который по своему химическому составу является нуклеотидом.
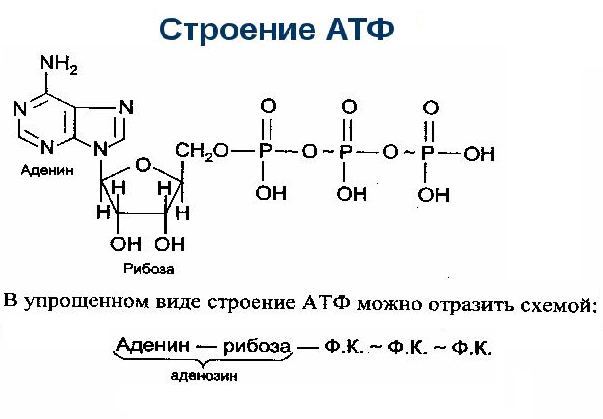
Молекула АТФ состоит из:
- – азотно-аденинового основания пурина;
- – пентозы, рибозы и моносахариды;
- – трех фосфатных групп, обозначенных как альфа, бета и гамма (начиная с рибозы).
Синтез АТФ в организме
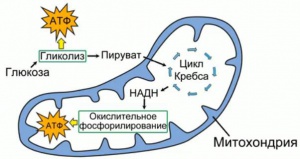
АТФ чаще всего производится в митохондрии, в основном в результате расщепления глюкозы и жирных кислот в процессе, называемом окислительным фосфорилированием; разложение 1 молекулы глюкозы в митохондрии высвобождает 36 молекул АТФ. Также АТФ синтезируется в хлоропластах, при фотосинтезе в процессе фотосинтетического фосфорилирования.
Использование АТФ в клетке
АТФ не может храниться в качестве резерва, поэтому он расходуется после его синтеза путем дефосфорилирования с помощью фермента АТФазы. Две конечные фосфорные группы связаны богатыми энергией ковалентными связями. Когда эти связи разрушаются, высвобождается относительно большое количество энергии. Если от АТФ освободить один конец ФГ, то образуется аденозин дифосфат (АДФ), освободить другой – получится аденозинмонофосфат (АМФ).
Фосфорная группа, высвобождаемая из АТФ или АДФ, богата энергией и, связываясь с соединением, обогащает ее энергией (процесс, называемый фосфорилированием). Таким образом, энергия от АТФ используется в процессах анаболизма.
АТФ создается в качестве основного энергетического продукта процесса разложения пищевых ингредиентов в процессе окисления. Часть энергии, выделяемой в этих процессах, сохраняется в форме АТФ, а остальная часть используется в форме тепла. Полученный таким образом АТФ используется для взаимодействия со всеми типами клеток. Только около 1/3 АТФ расходуется на реакции анаболизма. Остальная энергия расходуется на движение, сокращение мышц, транспортировку вещества через клеточную мембрану и т. д.
Фосфорилирование, регенерация АТФ.
Восстановление (синтез) АТФ реализуется путем связывания ФГ сначала с АМФ, что приводит к АДФ, а затем из АТФ под контролем фермента АТФ-синтазы. Это возможно благодаря тепловым реакциям, в которых энергоемкие (анаболические) реакции связаны с энерговыделительными (катаболическими) реакциями. Энергия, выделяемая при катаболизме, используется для повторного синтеза АТФ из АДФ. Следовательно, система АТФ / АДФ служит универсальным способом обмена энергией, который балансирует между выделяемыми и потребляющими энергию реакциями.
Функциональные характеристики АТФ.
Химическая связь, представляющая собой сумму сил, которые удерживают вместе атомы в молекуле, является стабильной конфигурацией, и для разрыва старой связи и образования новой требуется энергия. Ферменты значительно снижают потребность в активации большого количества энергии, но для того, чтобы химические реакции происходили в живых организмах, необходимо, чтобы энергия связи в продуктах реакции всегда была меньше энергии связи реагентов.
Молекула, наиболее часто участвующая в тепловых реакциях, – АТФ. Внутренняя структура молекул АТФ отлично подходит для этой роли в живых системах. В лабораторных условиях при удалении третьей фосфатной группы образуются АДФ и фосфат, и выделяется около 7 ккал (30 кДж) на моль АТФ. Удаление второй фосфатной группы дает AMФ и фосфат, высвобождая такое же количество энергии.
Энергия, выделяемая при удалении фосфатных групп, не только возникает из высокоэнергетических связей, но также является результатом перераспределения орбит в молекулах АТФ или АДФ. Каждая фосфатная группа несет отрицательный заряд и поэтому имеет тенденцию отталкиваться от другой такой группы. Когда фосфатная группа удаляется, происходит изменение конфигурации электронов, в результате чего получается структура с меньшей энергией.
В живых системах АТФ также гидролизуется до АДФ. Гидролиз АТФ является, например, быстрым способом выработки тепла у животных, которые просыпаются от зимней спячки. Однако обычно конечный продукт не просто удаляется, а переносится через фермент (киназу) в другую молекулу (фосфорилирование). Эта реакция также передает часть энергии от высокоэнергетической связи фосфорилированному соединению, которое, таким образом, обогащается энергией при реакции.
Энергия, выделяемая в реакциях клеточного метаболизма, таких как расщепление глюкозы, используется для повторного синтеза АТФ из молекул АДФ. Основными механизмами синтеза АТФ в клетке являются окислительное фосфорилирование в процессе клеточного дыхания (на внутренней стороне митохондриальной мембраны) и фосфорилирование в процессе фотосинтеза.
Митохондрии
Митохондрии представляют собой мембранные органеллы, присутствующие в клетках практически всех эукариотических организмов. Митохондрии заключены в две мембраны: внешняя, находящаяся в контакте с цитоплазмой, и мембрана, ограничивающая внутреннюю часть митохондрий. Между этими двумя мембранами находится межмембранное пространство. Внутренняя часть митохондрий заполнена матриксом. Типичная эукариотическая клетка содержит около 2000 митохондрий.
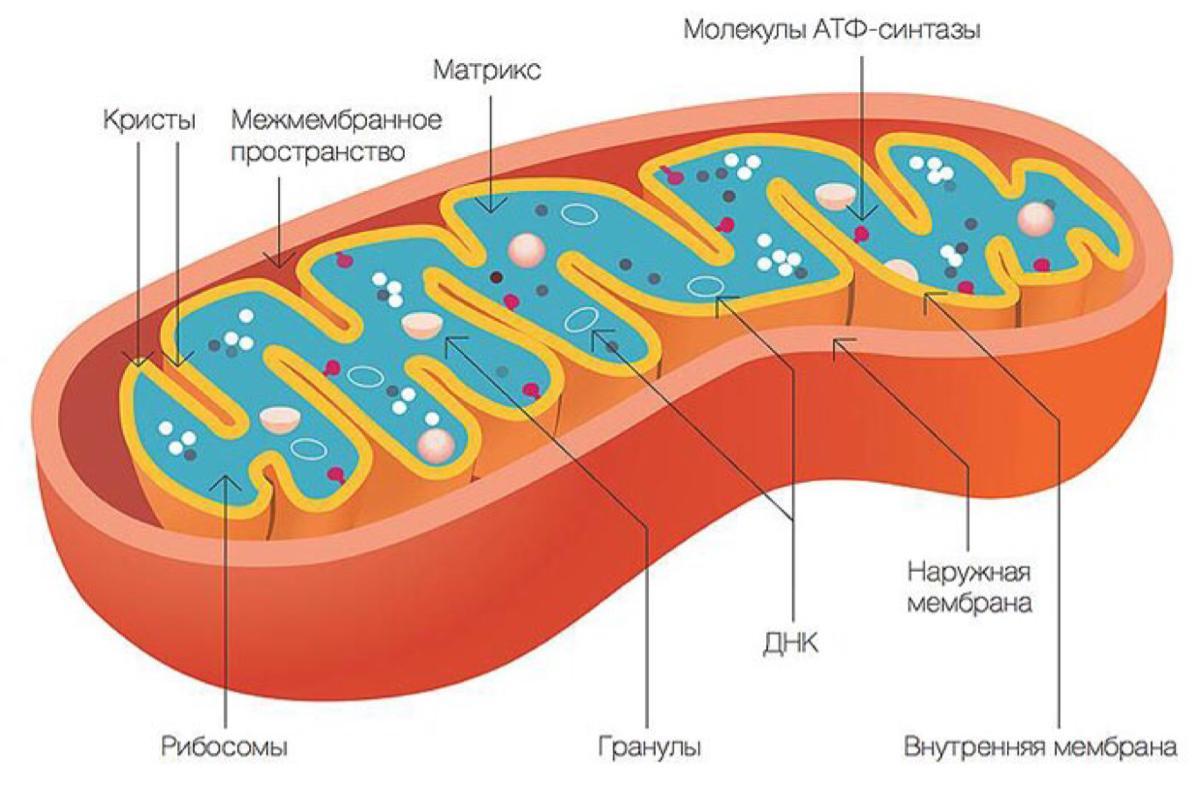
Внешняя митохондриальная мембрана определяет форму этой органеллы и, благодаря наличию каналообразующего белка (порина), проницаема для определенных молекул.
Внутренняя митохондриальная мембрана имеет в несколько раз большую площадь поверхности, чем наружная мембрана. Ее поверхность значительно увеличивают кристы, ориентированные к центру органеллы. Кристы могут различаться по количеству, размеру и форме, при этом они имеют частицы, прикрепленные к ним с помощью коротких ручек. Эти частицы содержат АТФ-синтазы, ферментный комплекс, участвующий в синтезе АТФ.
Матрикс заполняет внутреннюю часть митохондрий и представляет собой смесь нескольких сотен ферментов, которые преобразуют продукты метаболизма углеводов, липидов и белков через цикл Кребса в углекислый газ и воду с выделением энергии в виде молекул АТФ. В этом процессе электроны переносятся по дыхательной электронной цепи, и происходит синтез высокоэнергетического фосфатного соединения, АТФ (окислительное фосфорилирование).
Источник

Общепринятая на данный момент теория симбиогенеза предполагает, что митохондрии в эукариотических клетках произошли от симбиотических бактерий. Однако поиски предковой бактерии и реконструкция событий симбиогенеза еще далеки от завершения. Авторы новой статьи в журнале PNAS подошли к проблеме с другого конца: они смоделировали симбиогенез на примере хорошо изученной бактерии (Escherichia coli) и хорошо изученной эукариотической клетки (Saccharomyces cerevisiae). Теперь у нас есть отработанная методика получения химерных клеток, с помощью которой можно проверять, какие именно свойства предковой бактерии были необходимы для симбиогенеза.
Теория симбиогенеза была предложена в 1967 году. Согласно ей, археи и протеобактерии вступили в эндосимбиоз (первые тем или иным способом «поглотили» вторых), что привело к возникновению эукариот (см.: Теория симбиогенеза 50 лет спустя: параллельной эукариотизации, скорее всего, не было, «Элементы», 22.11.2017). За 50 лет удалось уточнить разные детали: судя по всему, эндосимбиоз с предками митохондрий произошел только один раз, а не в нескольких параллельных ветвях, и это стало конечным этапом в становлении эукариот (см.: Генеалогия белков свидетельствует о позднем приобретении митохондрий предками эукариот, «Элементы», 08.02.2016). Однако вопросов всё еще остается немало: например, что это была за предковая бактерия? Одна из распространенных точек зрения заключается в том, что изначально бактерии паразитировали на клетках архей, а потом паразитизм перешел в симбиоз. В таком случае, ближайшие родственники такой бактерии, известные нам, – это альфапротеобактерии риккетсии, внутриклеточные паразиты многих животных и человека (вызывают, например, эпидемический сыпной тиф и пятнистую лихорадку Скалистых гор).
Можно продолжать поиск родственников «с конца», то есть сравнивать геномы современных митохондрий с геномами различных бактерий и искать пересечения, а можно зайти «с начала» и попробовать воспроизвести эту предковую бактерию самим. Для этого нужно определить минимальный набор свойств, которыми она должна обладать для успешного внедрения внутрь археи. Заодно такой метод мог бы пролить свет на последовательность событий симбиогенеза. Но коль скоро мы не умеем создавать бактерии с нуля, можно модифицировать самую изученную бактерию на свете – кишечную палочку (Escherichia coli).
Общий принцип, которым руководствовались авторы эксперимента, можно сформулировать так: чтобы заставить две клетки вступить в симбиоз, нужно отобрать у них что-то жизненно важное, тогда их существование по отдельности станет невозможно (рис. 2).

Всю работу можно условно разделить на пять шагов.
Шаг 1 – лишить кишечную палочку самодостаточности. Чтобы эндосимбиоз оказался выгодным решением для бактерии, она должна стать ауксотрофом – быть неспособной производить какое-нибудь жизненно необходимое вещество. Для многих бактерий таким веществом является тиамин (витамин B1) – кофермент в реакциях углеводного обмена. Поэтому в геноме E. coli ген биосинтеза тиамина был заменен на кассету (см. Gene cassette) с GFP (зеленым флуоресцентным белком) и геном устойчивости к антибиотику канамицину. Теперь клетки не могут выживать без внешнего источника тиамина (который они сквозь мембрану закачивают внутрь), их можно отобрать под действием антибиотика и отследить во флуоресцентный микроскоп.
Шаг 2 – сделать кишечную палочку полезной. Авторы гипотезы происхождения митохондрии из внутриклеточных паразитов полагают, что одним из ключевых белков был АТФ/АДФ-антипортер (см. Antiporter). Это белок-переносчик, который обменивает АТФ на АДФ, меняя их местами по разные стороны мембраны. У паразитической бактерии он должен работать на благо бактерии: захватывать АТФ снаружи (то есть отбирать у клетки-хозяина) и менять на отработанные АДФ бактерии. Однако этот механизм можно заставить работать и в обратную сторону, если концентрации веществ поменяются местами. При этом бактерия начнет забирать АДФ из цитоплазмы хозяина и отдавать АТФ. Так или иначе, АДФ/АТФ-антипортеры есть как у современных митохондрий, так и у внутриклеточных паразитов. У свободно живущей кишечной палочки такого белка нет, поэтому пришлось снабдить клетки E. coli плазмидой с соответствующим геном.
Шаг 3 – лишить дрожжи самодостаточности. Чтобы заставить дрожжи вступить в симбиоз, их нужно лишить энергии, то есть АТФ. Тогда единственным выходом будет получить его от кишечной палочки. Но у дрожжей, как у почти всех эукариот, есть свои митохондрии. Поэтому авторы эксперимента взяли мутантный штамм дрожжей, лишенный одного из ключевых митохондриальных генов. Такие клетки содержат митохондрии, но не получают от них энергии. Они не могут расти в среде, где из питательных веществ есть только глицерин. Однако оказалось, что и в симбиоз с E. coli они тоже не вступают.
Шаг 4 – добавить «белки слияния». Эукариотическая клетка – это множество вложенных друг в друга мембранных пузырьков. Чтобы органеллы хаотично не сливались друг с другом, мембраны покрыты белками группы SNARЕ, которые могут стимулировать или блокировать слияние. Многие патогенные бактерии тоже несут SNARE-подобные белки. Клетка-хозяин воспринимает их как собственные органеллы и не переваривает (то есть с ними не сливаются лизосомы). Правда, мы пока не уверены в том, что к моменту эндосимбиоза эукариоты уже обладали системой этих белков. Но коль скоро мы работаем с дрожжами, приходится на нее ориентироваться. Авторы эксперимента ввели кишечной палочке гены трех разных SNARE-подобных белков, позаимствованных у хламидий. И только после этого они получили устойчивые колонии дрожжей с симбиотическими E. coli (рис. 3). Колонии росли на среде, богатой глицерином, лишенной тиамина и с добавлением антибиотика канамицина, – то есть удовлетворяли всем условиям эксперимента. В том же составе химерные клетки размножались в течение последующих трех дней культивирования, что соответствует примерно 40 делениям.

Шаг 5 – убрать лишнее. В ходе эволюции митохондрия утратила большую часть ДНК (у млекопитающих, например, в ее геноме осталось лишь 37 генов). Это значит, что она становилась всё более зависимой от своей клетки-хозяина. Авторы обсуждаемой статьи попробовали воспроизвести и этот этап тоже. Для этого они удалили у клеток кишечной палочки ген биосинтеза НАД+ – еще одного важного кофермента. Клетки, лишенные НАД+, так же как и их предшественники, лишенные тиамина, успешно образовывали химеры с дрожжами. И даже двойные мутанты, неспособные производить ни один из этих коферментов, также вступали в эндосимбиоз (рис. 4).

Перед нами – отработанная методика, с помощью которой можно моделировать ранние события эндосимбиоза. Клетки кишечной палочки, дефицитные по разным веществам, равно хорошо образуют химеры, которые воспроизводятся из поколения в поколение. Следующий шаг – поиск предельной редукции генома E. coli, возможной в данной ситуации. Авторы статьи отмечают, что удаление всего двух путей биосинтеза уже дало экономию в 7,7 тысяч пар нуклеотидов (для сравнения, весь митохондриальный геном человека составляет примерно 15 тысяч пар). Поэтому нам еще предстоит найти ту грань, на которой экономия размера генома столкнется с возможностью выживания клетки-симбионта.
Кроме того, как ехидно указывают авторы в конце текста, при таком раскладе не очень понятно, кто в этой истории настоящий паразит. Если бактерия, попавшая внутрь археи, лишь постепенно утрачивала свои метаболические пути, то возможно настоящим паразитом здесь стоит считать архею, которая потребляла энергию, производимую бактерией.
Источник: A. P. Mehta, L. Supekova, J.-H. Chen, K. Pestonjamasp, P. Webster, Y. Ko, S. C. Henderson, G. McDermott, F. Supeke, P. G. Schultz. Engineering yeast endosymbionts as a step toward the evolution of mitochondria // PNAS. a of October 29, 2018. DOI: 10.1073/pnas.1813143115.
Полина Лосева
Источник