Для регуляции биосиотеза аминокислот кишечной палочкой характерно

1)гидролиз природного белковосодержащего сырья;
2)химический синтез с разделением рацематов на иммобилизованной аминоацилазе
3)химико-ферментативный синтез
4)микробиологический синтез
300.МЕХАНИЗМ КОНТРОЛЯ СКОРОСТИ БИОСИНТЕЗА АМИНОКИСЛОТЫ У ПРИРОДНОГО ПРОДУЦЕНТА – КИШЕЧНОЙ ПАЛОЧКИ, ПРЕПЯТСТВУЮЩИЙ ИЗБЫТОЧНОМУ НАКОПЛЕНИЮ АМИНОКИСЛОТЫ
1)не согласованная репрессия
2)согласованная репрессия
3)совместное ингибиторование
4)репрессия
301.У ТИПИЧНЫХ ПРИРОДНЫХ НЕ МУТАНТНЫХ ПРОДУЦЕНТОВ
ЛИЗИНА Согуnebacterium glutamicum И, Brevibacterium flavum ФЕРМЕНТ АСПАРТАТКИНАЗА ЯВЛЯЕТСЯ АЛЛОСТЕРИЧЕСКИМ БЕЛКОМ, ЧУВСТВИТЕЛЬНЫМ ПО ПРИНЦИПУ ОБРАТНОЙ СВЯЗИ ПРИ СОВМЕСТНОМ ДЕЙСТВИИ
1)только лизина
2)только треонина
3)L- лизина и L- треонина
4)D- лизина и L- лизина
302.КАКОЙ ИЗ ПРИМЕНЯЕМЫХ МЕТОДОВ ПРОМЫШЛЕННОГО
ПОЛУЧЕНИЯ АМИНОКИСЛОТ ЯВЛЯЕТСЯ ПОЛНОСТЬЮ БИОТЕХНОЛОГИЧЕСКИМ (БАЗИРУЕТСЯ ЦЕЛИКОМ НА ПРИМЕНЕНИИ БИООБЪЕКТОВ)
1)гидролиз природного белковосодержащего сырья;
2)химический синтез с разделением рацематов на иммобилизованной аминоацилазе
3)химико-ферментативный синтез
4)микробиологический синтез
303.Corinebacterium glutamicum ЯВЛЯЕТСЯ ПРОДУЦЕНТОМ ДЛЯ СЛЕДУЮЩИЕЙ АМИНОКИСЛОТЫ
1)треонин
2)триптофан
3)фенилаланин
4)лейцин
304.Corinebacterium glutamicum ЯВЛЯЕТСЯ ПРОДУЦЕНТОМ ДЛЯ СЛЕДУЮЩИЕЙ АМИНОКИСЛОТЫ
1)лейцин
51
2)гистидин
3)изолейцин
4)валин
305.Corinebacterium glutamicum ЯВЛЯЕТСЯ ПРОДУЦЕНТОМ ДЛЯ СЛЕДУЮЩИЕЙ АМИНОКИСЛОТЫ
1)серин
2)Фенилаланин
3)изолейцин
4)триптофан
306.ДЛЯ РЕГУЛЯЦИИ БИОСИНТЕЗА АМИНОКИСЛОТ У КОРИНЕБАКТЕРИЙ ХАРАКТЕРНО
1)ретроингибирование
2)согласованная репрессия
3)совместное ингибирование
4)ауксотрофен
307.СИНТЕЗ ЛИЗИНА ОСУЩЕСТВЛЯЮТ КОРИНЕБАКТЕРИИ, АУКСОТРОФНЫЕ ПО
1)изолейцину
2)треонину
3)лизину
4)валину
308.СИНТЕЗ ЛИЗИНА ОСУЩЕСТВЛЯЮТ КОРИНЕБАКТЕРИИ, АУКСОТРОФНЫЕ ПО
1)изолейцину
2)лизину
3)гомосерину
4)валину
309.АМИНОКИСЛОТУ ТРЕОНИН ПРОДУЦИРУЮТ МУТАНТНОИНЖЕНЕРНЫЕ ШТАММЫ
1)стрептококков
2)кишечной палочки
3)коринебактерий
4)пекарских дрожжей
310.МУТАНТНО-ИНЖЕНЕРНЫЙ ШТАММ КИШЕЧНОЙ ПАЛОЧКИ – ПРОДУЦЕНТ ТРЕОНИНА
1)ауксотрофен по тренину и гомосерину
2)синтезирует продукт после накопления биомассы
3)не нуждается в аминокислотах для своего роста
4)синтезирует продукт до накопления биомассы
52
311.ДЛЯ РЕГУЛЯЦИИ БИОСИНТЕЗА АМИНОКИСЛОТ КИШЕЧНОЙ ПАЛОЧКОЙ ХАРАКТЕРНО
1)репрессия
2)ретроингибирование
3)совместное ингибирование лизином и треонином
4)согласованная репрессия треонином и изолейцином
312.АМИНОКИСЛОТУ ЛИЗИН ПРОДУЦИРУЮТ МУТАНТНЫЕ
ШТАММЫ
1)кишечной палочки
2)коринебактерий
3)пекарских дрожжей
4)стрептококков
313.РЕЗИДЕНТНОЙ НАЗЫВАЮТ
1)условно-патогенную микрофлору ЖКТ
2)патогенную микрофлору ЖКТ
3)постоянную микрофлору ЖКТ
4)транзиторную микрофлору ЖКТ
314.КУЛЬТИВИРОВАНИЕ КИШЕЧНОЙ ПАЛОЧКИ ОСУЩЕСТВЛЯЮТ, ПРИ РН
1)рН = 5,5-6,0
2)рН = 8,0-8,2
3)рН = 6,0-7,0
4)рН = 7,2-8,0
315.К ПРЕПАРАТАМ ПРОБИОТИКОВ, НЕ СОДЕРЖАЩИМ БИФИДОБАКТЕРИИ, ОТНОСЯТ
1)пробифор
2)нормофлор
3)бификол
4)бифилиз
316.К ПРЕПАРАТАМ ПРОБИОТИКОВ, НЕ СОДЕРЖАЩИЕ ЛАКТОБАКТЕРИИ, ОТНОСЯТ
1)гастрофарм
2)бифилиз
3)линекс
4)лактобактерин сухой
317.ЕСЛИ ОБА ШТАММА В СМЕШАННОЙ КУЛЬТУРЕ РАСТУТ БЫСТРЕЕ, ЧЕМ В СООТВЕТСТВУЮЩИХ ЧИСТЫХ КУЛЬТУРАХ, ЯВЛЕНИЕ НОСИТ НАЗВАНИЕ
53
1)нейтрализм
2)мутуализм
3)аменсализм
4)комменсализм
318.РОСТ ОДНОГО МИКРООРГАНИЗМА ПОДАВЛЯЕТСЯ В ПРИСУТСТВИИ ДРУГОГО — ЭТО
1)нейтрализм
2)аменсализм
3)комменсализм
4)симбиоз
319.КУЛЬТИВИРОВАНИЕ МОЛОЧНО-КИСЛЫХ БАКТЕРИЙ ОСУЩЕСТВЛЯЮТ, ПРИ РН
1)рН = 5,5-6,0
2)рН = 8,0-8,2
3)рН = 6,0-7,0
4)рН = 7,2-8,0
320.МЕХАНИЗМЫ МУТУАЛИЗМА
1)обмен питательными веществами
2)синтез токсических веществ
3)поглощение незаменимых питательных веществ
4)секреция ферментов, разрушающих полимеры клеточной стенки
321.СОСТОЯНИЕ СИСТЕМЫ, КОГДА НИ ОДИН ИЗ ОРГАНИЗМОВ НЕ ОКАЗЫВАЕТ ВЛИЯНИЯ НА СКОРОСТЬ РОСТА ДРУГОГО МИКРООРГАНИЗМА, НАЗЫВАЕТСЯ
1)нейтрализм
2)мутуализм
3)комменсализм
4)аменсализм
322.РОСТ ОДНОГО МИКРООРГАНИЗМА ПОДАВЛЯЕТСЯ В ПРИСУТСТВИИ ДРУГОГО – ЭТО
1)нейтрализм
2)аменсализм
3)комменсализм
4)симбиоз
323.ЕСЛИ В СМЕШАННОЙ КУЛЬТУРЕ ПРЕИМУЩЕСТВА ПОЛУЧАЕТ ВТОРОЙ ВИД МИКРООРГАНИЗМОВ, ТО ЯВЛЕНИЕ НАЗЫВАЮТ
1)аменсализм
54
2)мутуализм
3)комменсализм
4)симбиоз
324.ПАРАЗИТИЗМОМ НАЗЫВАЮТ ВАРИАНТ
1)мутуализма
2)аменсализма
3)комменсализма
4)симбиоз
325.МЕТАБОЛИЗМ ХОЛЕСТЕРИНА ОСУЩЕСТВЛЯЮТ
1)бифидобактерии
2)лактобактерии
3)непатогенные штаммы кишечной палочки
4)грибы рода Кандида
326.СИМБИОЗОМ НАЗЫВАЮТ
1)тесные мутуалистические связи
2)тесные аменсалитические связи
3)тесные комменсалитические связи
4)аменсализм
327.ДИАРЕЯ ПУТЕШЕСТВЕННИКОВ ОБУСЛОВЛЕНА
1)снижением количества бифидо- и лактобактерий
2)развитием кишечных палочек с патогенными свойствами
3)развитием дрожжеподобных грибов рода Кандида
4)постоянной микрофлорой ЖКТ
328.НАКОПЛЕНИЕ БИОМАССЫ КУЛЬТУРЫ КИШЕЧНОЙ ПАЛОЧКИ ПРОВОДЯТ НА ПИТАТЕЛЬНЫХ СРЕДАХ НА ОСНОВЕ
1)казеина и желатина
2)печеночного бульона, пептона и лактозы
3)гидролизата молока, солодового экстракта, глюкозы
4)мелассы и хлорида натрия
329.В КАЧЕСТВЕ ЗАЩИТНОЙ СРЕДЫ ПРИ ЛИОФИЛЬНОЙ СУШКЕ СУСПЕНЗИИ КИШЕЧНОЙ ПАЛОЧКИ В ПРОИЗВОДСТВЕ КОЛИБАКТЕРИНА ИСПОЛЬЗУЮТ
1)сахарозу
2)глюкозу
3)пептон
4)обрат молока
330.СИМБИОНТАМИ МАКРООРГАНИЗМА С ПЕРВЫХ ДНЕЙ ЖИЗНИ ЯВЛЯЮТСЯ
55
1)бифидобактерии
2)кишечная палочка
3)бактероиды
4)грибы рода Кандида
331.Bacillus ВХОДИТ В СОСТАВ ПРЕПАРАТА
1)Флонивин БС
2)Нормофлор
3)Энтерол
4)Бификол
332.В КАЧЕСТВЕ ЗАЩИТНОЙ СРЕДЫ ПРИ ЛИОФИЛЬНОЙ СУШКЕ СУСПЕНЗИИ БИФИДОБАКТЕРИЙ В ПРОИЗВОДСТВЕ БИФИДОБАКТЕРИНА ИСПОЛЬЗУЮТ
1)сахарозу
2)глюкозу
3)пептон
4)обезжиренное молоко
333.ПРЕПАРАТ НОРМОФЛОР СОДЕРЖИТ ЛИОФИЛИЗИРОВАННЫЕ
КЛЕТКИ
1)Bacillus subtilis
2)Lactobaccillus acidophilus
3)Lactobaccillus bulgaricus
4)Kefir greins
334.НАКОПЛЕНИЕ БИОМАССЫ КУЛЬТУР LACTOBACILLUS ПРОВОДЯТ НА ПИТАТЕЛЬНЫХ СРЕДАХ НА ОСНОВЕ
1)казеина и желатина
2)печеночного бульона, пептона и лактозы
3)гидролизата молока, солодового экстракта, глюкозы
4)мелассы и хлорида натрия
335.ВЫВОДЯТСЯ ИЗ ОРГАНИЗМА ПОСЛЕ КУРСА ЛЕЧЕНИЯ ПРОБИОТИКИ, ВХОДЯЩИЕ В СОСТАВ ПРЕПАРАТОВ
1)Бифилиз
2)Энтерол
3)Бификол
4)Колибактерин
336.ПРЕПАРАТЫ ПРОБИОТИКОВ, СОДЕРЖАЩИЕ НЕСКОЛЬКО ШТАММОВ МИКРООРГАНИЗМОВ
1)Гастрофарм
2)Линекс
3)Энтерол
56
4)Бифилиз
337.ЛИЗОЦИМ ВХОДИТ В СОСТАВ ПРЕПАРАТА
1)Флонивин БС
2)Бактисубтил
3)Бифилиз
4)Бификол
338.ОСНОВНОЕ ПРЕИМУЩЕСТВО ФЕРМЕНТАТИВНОЙ БИОКОНВЕРСИИ СТЕРОИДОВ ПЕРЕД ХИМИЧЕСКОЙ ТРАНСФОРМАЦИЕЙ СОСТОИТ
1)в доступности реагентов
2)в избирательности воздействия на определенные функциональные группы стероида
3)в сокращении времени процесса
4)в получении принципиально новых соединений
339.ПРЕИМУЩЕСТВОМ МЕТОДА БИОКОНВЕРСИИ СТЕРОИДОВ ПЕРЕД ХИМИЧЕСКОЙ ТРАНСФОРМАЦИЕЙ ЯВЛЯЕТСЯ
1)высокая скорость реакции окисления
2)окисление только по боковой цепи
3)окисление по системе сконденсированных колец
4)окисление как по системе колец, так и по боковой цепи
340.ВЕЩЕСТВО S РАЙХШТЕЙНА МОЖЕТ БЫТЬ ПОЛУЧЕНО ИЗ
1)аланина
2)соласодина
3)преднизолона
4)целлюлозы
341.КОРТИКОСТЕРОИДЫ СОДЕРЖАТ ПРИ С-17
1)аминогруппу
2)гидроксизамещенную ацетильную группу
3)кольцо ароматическое
4)карбонильную или гидроксильную группы, а их модифицированные аналоги — алкильную или этинильную группу
342.УВЕЛИЧЕНИЕ ВЫХОДА ЦЕЛЕВОГО ПРОДУКТА ПРИ БИОТРАНСФОРМАЦИИ СТЕРОИДА ДОСТИГАЕТСЯ
1)При увеличении интенсивности перемешивания
2)при увеличении интенсивности аэрации
3)при повышении температуры ферментации
4)при увеличении концентрации стероидного субстрата в ферментационной среде
57
343.ВЕЩЕСТВО S РАЙХШТЕЙНА МОЖЕТ БЫТЬ ПОЛУЧЕНО ИЗ
1)диосгенина
2)аланина
3)преднизолона
4)целлюлозы
344.МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПОЛУЧАЮТ В ПРОИЗВОДСТВЕ
1)при фракционировании антител организмов
2)фракционированием лимфоцитов
3)с помощью гибридом
4)химическим синтезом
345.ПО ПРОИСХОЖДЕНИЮ ИММУНОСТИМУЛЯТОРЫ ПОДРАЗДЕЛЯЮТ НА
1)экзогенные
2)химические
3)биосинтетические
4)экстракционные
346.ЭНДОГЕННЫЕ ИММУНОСТИМУЛЯТОРЫ СИНТЕЗИРУЮТСЯ 1)клетками микроорганизмов
2)с помощью химических реакций
3)клетками макроорганизма
4)половыми клетками
347.ELISA — ТВЕРДОФАЗНЫЙ ИММУНОФЕРМЕНТНЫЙ АНАЛИЗ ЯВЛЯЕТСЯ
1)иммунометрическим
2)конкурентным
3)быстрым
4)гетерогенным
348.«СЕНДВИЧ» – АНАЛИЗ ЯВЛЯЕТСЯ
1)иммунометрическим
2)конкурентным
3)гомогенным
4)гетерогенным
349.ВАРИАНТЫ ПОСТАНОВКИ ИФА
1)онкурентный, иммунометрический
2)юминисцентным
3)адиоиммунный
4)люоресцертный
58
350.ВАКЦИНЫ ФОРМИРУЮТ ИММУНИТЕТ
1)пассивный
2)активный
3)быстрый
4)медленный
351.ПРЕИМУЩЕСТВА ИФА ПЕРЕД МЕТОДОМ РИА
1)меньшая стоимость анализа
2)легкость освоения персоналом
3)отсутствие радиоактивных изотопов
4)возможность визуальной оценки результата
352.«СЕНДВИЧ» – АНАЛИЗ ПРИМЕНИМ
1)к поликлональным иммуноглобулинам
2)к ионоклональным антителам
3)как к поли так и к моноклональным антителам
4)к аминокислотам
353.ТОЧНОСТЬ ОПРЕДЕЛЕНИЯ ВЫШЕ В МЕТОДЕ
1)ELISA
2)«СЕНДВИЧ»
3)EMIT
4)РИА
354.В КАЧЕСТВЕ МАРКЕРА В ТЕСТЕ ИФА УСТАНОВЛЕНИЯ ФАКТА БЕРЕМЕННОСТИ ИСПОЛЬЗУЮТ
1)йод-125
2)тритий
3)пероксидазу
4)галактозидазу
355.ГОМОГЕННЫЙ ИФА ОСНОВАН
1)на разделении компонентов после проведения реакции
2)на изменении активности фермента в процессе реакции
3)на адсорбции фермента на носителе
4)на подавление фермента
356.ПО ПРОИСХОЖДЕНИЮ ИММУНОСТИМУЛЯТОРЫ ПОДРАЗДЕЛЯЮТ НА
1)эндогенные
2)экзогенные
3)химические
4)биосинтетические
59
357.ПО ПРОИСХОЖДЕНИЮ ИММУНОСТИМУЛЯТОРЫ ПОДРАЗДЕЛЯЮТ НА
1)эндогенные
2)экстракционные
3)химические
4)биосинтетические
358.ELISA — ТВЕРДОФАЗНЫЙ ИММУНОФЕРМЕНТНЫЙ АНАЛИЗ ЯВЛЯЕТСЯ
1)иммунометрическим
2)конкурентным
3)гомогенным
4)быстрым
359.АКТИВНОСТЬ АЛЬФА-ИНТЕРФЕРОНА ОПРЕДЕЛЯЕТСЯ ПО ЗАЩИТНОМУ ПРОТИВОВИРУСНОМУ ДЕЙСТВИЮ НА КУЛЬТУРУ КЛЕТОК
1)яичников китайского хомячка
2)эмбрионов человека
3)печени обезьяны
4)куринной эмбриональной ткани
360.В ПРОИЗВОДСТВЕ РЕКОМБИНАНТНЫХ В- И У- ИНТЕРФЕРОНОВ ИСПОЛЬЗУЮТ ЭУКАРИОТНЫЕ ПРОДУЦЕНТЫ БЛАГОДАРЯ ИХ СПОСОБНОСТИ ОСУЩЕСТВЛЯТЬ
1)сплайсинг
2)процессинг
3)продуцирование внеклеточных метаболитов
4)гликозилирование белков
361.ВЫДЕЛЕНИЕ И ОЧИСТКУ ИНТЕРФЕРОНОВ ПРОВОДЯТ МЕТОДОМ
1)гель-хроматографии
2)аффинной хроматографии
3)ионнообменной хроматографии
4)адсорбционной хроматографии
362.ПРЕПАРАТЫ РЕКОМБИНАНТНОГО АЛЬФА-ИНТЕРФЕРОНА
1)виферон
2)эгиферон
3)циклоферон
4)линекс
363.РАЗРАБОТАННЫЕ ТЕХНОЛОГИИ РЕКОМБИНАНТНОГО Α- ИНТЕРФЕРОНА ОСНОВАНЫ НА ЭКСПРЕССИИ ГЕНА
60
Источник
a)□ | оксидо-редуктаз |
b)□ | лигаз |
c)□ | изомераз |
d)□ | лиаз |
e)□ | гидролаз |
f)□ | трансфераз |
При культивировании дрожжеподобных грибов рода Candida можно получить:
a)□ | эргостерин и рибофлавин |
b)□ | убихинон и эргостерин |
c)□ | эргостерин и витамин В 12 |
d)□ | убихинон и бета-каротин |
При культивировании уксуснокислых бактерий можно получить:
a)□ | L-сорбозу и убихинон |
b)□ | L-сорбозу и рибофлавин |
c)□ | витамин С и бета-каротин |
d)□ | L-сорбозу и эргостерин |
Убихиноны участвуют в биохимических реакциях:
a)□ | окисления жирных кислот |
b)□ | восстановления |
c)□ | окислительного фосфорилирования |
d)□ | декарбоксилирования |
Гидролиз L-изомеров ацилированных аминокислот осуществляет иммобилизованный фермент:
a)□ | аминорацемаза |
b)□ | аминоизомераза |
c)□ | аминоацилаза |
d)□ | аминотрансфераза |
Химико-ферментативный синтез аспарагиновой кислоты из фумаровой кислоты в присутствии аммиака осуществляют:
a)□ | клетки коринебактерий |
b)□ | клетки кишечной палочки |
c)□ | иммобилизованные клетки кишечной палочки |
d)□ | иммобилизованные клетки дрожжей |
Аминокислоту треонин продуцируют мутантно-инженерные штаммы:
a)□ | коринебактерий |
b)□ | стрептококков |
c)□ | пекарских дрожжей |
d)□ | кишечной палочки |
Для регуляции биосинтеза аминокислот кишечной палочкой характерно:
a)□ | совместное ингибирование лизином и треонином |
b)□ | согласованная репрессия треонином и изолейцином |
c)□ | ретроингибирование |
d)□ | репрессия |
Аминокислоту лизин продуцируют мутантные штаммы:
a)□ | пекарских дрожжей |
b)□ | кишечной палочки |
c)□ | стрептококков |
d)□ | коринебактерий |
Для регуляции биосинтеза аминокислот у коринебактерий характерно:
a)□ | совместное ингибирование |
b)□ | согласованная репрессия |
c)□ | ретроингибирование |
Химико-ферментативный синтез фенилаланина из коричной кислоты и аммиака осуществляют иммобилизованные клетки:
a)□ | пекарских дрожжей |
b)□ | стрептококков |
c)□ | кишечной палочки |
d)□ | коринебактерий |
Промышленным продуцентом глутаминовой кислоты являются штаммы:
a)□ | Corynebacterium glutamicum |
b)□ | Penicillium solitum |
c)□ | Escherichia coli |
d)□ | Bacillus subtilis |
Биосинтез вторичных метаболитов фазоспецифичен и происходит в:
a)□ | фазе отмирания |
b)□ | логарифмической фазе |
c)□ | фазе замедления |
d)□ | лаг-фазе |
e)□ | фазе ускорения |
f)□ | стационарной фазе |
По способу культивирования и потребности в аэрации биотрансформация стероидов – это:
a)□ | анаэробный процесс поверхностной ферментации |
b)□ | аэробный процесс глубинной ферментации |
c)□ | анаэробный процесс глубинной ферментации |
Производство стероидного препарата преднизолона из кортикостерона осуществляется путем:
a)□ | микробного ацетилирования |
b)□ | микробного гидроксилирования |
c)□ | химического аминирования |
d)□ | химического ацетилирования |
Назовите микроорганизм, переводящий кортизол в преднизолон
a)□ | Aspergillus oryzae |
b)□ | Arthrobacter simplex |
c)□ | Curvularia lunata |
d)□ | Streptomyces albus |
Какое вещество является предшественником кортизола при синтезе стероидов?
a)□ | прогестерон |
b)□ | преднизолон |
c)□ | вещество S Рейхштейна |
d)□ | диосгенин |
Из желчных камней в 1782 г. был впервые выделен:
Источник
Большинство микроорганизмов, подобно высшим растениям, способны синтезировать все аминокислоты, входящие в состав клеточных белков. Исходным материалом для построения углеродных скелетов аминокислот служат промежуточные продукты обмена углеводов в реакциях гликолиза и цикла трикарбоновых кислот. Наиболее важными из них являются кетокислоты. Перевод азота в органические соединения осуществляется через аммиак. Нитраты, нитриты, молекулярный азот восстанавливаются до аммиака в результате ассимиляционной нитратредукции и включаются в состав органических соединений.
Некоторые аминокислоты образуются путем прямого аминировакия кетокислот аммонием. Аминогруппы вводятся путем прямого аминирования или транса минирования. Таким путем
происходит первичная ассимиляция аммиака и построение первичных аминокислот, которые служат источником аминных групп для образования других аминокислот путем переаминирования, т. е. переноса аминогрупп с аминокислот на кетокислоты. В реакциях прямого аминирования принимают участие глутаматдегидрогеназа и аланиндегидрогеназа. Так, а- кетоглутаровая кислота при участии глутаматдегидрогеназы реагирует с аммонием, образуя глутаминовую кислоту, аланин- дегидрогеназа катализирует включение аммония в пиро- виноградную кислоту и образование аланина; аспартатаммиаклиаза катализирует синтез аспарагиновой аминокислоты:
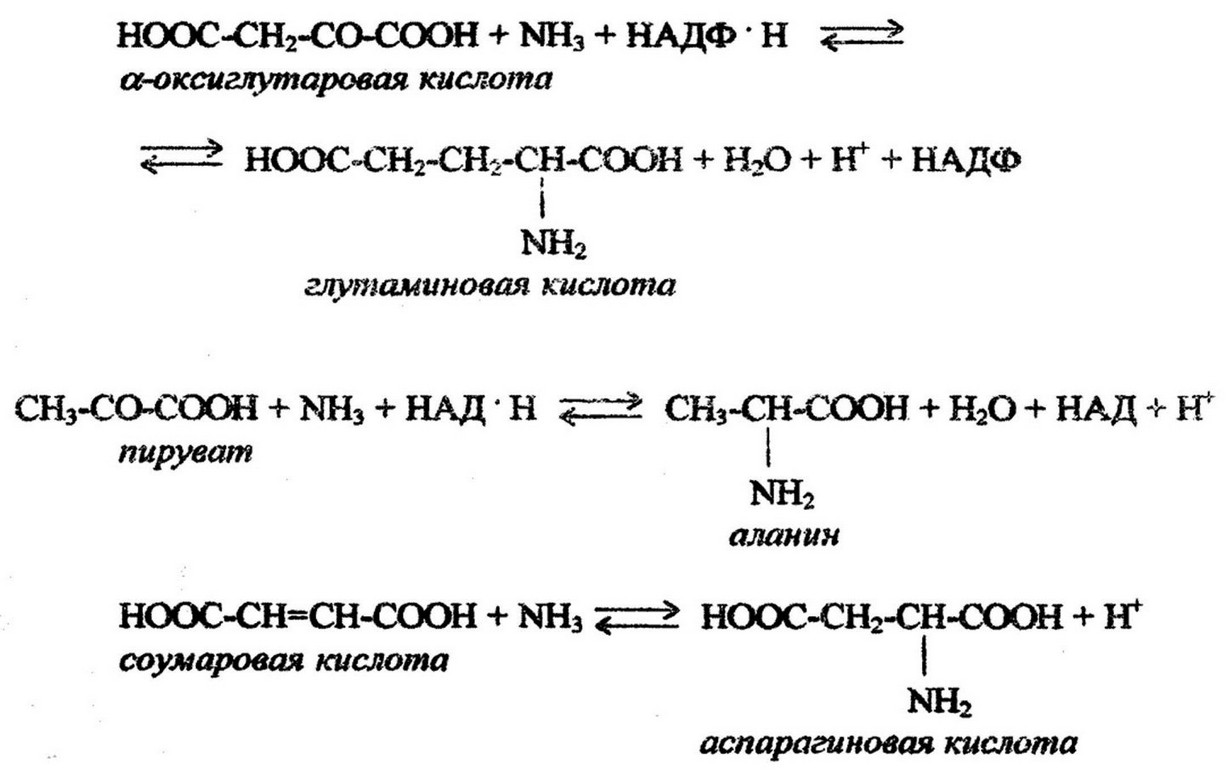
Многие гетеротрофные бактерии образуют новые аминокислоты в основном через трансаминирование или переаминирование аминокислот, потребляемых из питательной среды. Важным в биосинтетических реакциях является превращение одних аминокислот в другие: например, глутамата в аргинин и пролин; аспарагиновой кислоты в лизин, метионин, треонин; треонина в глюкокол и т. д.
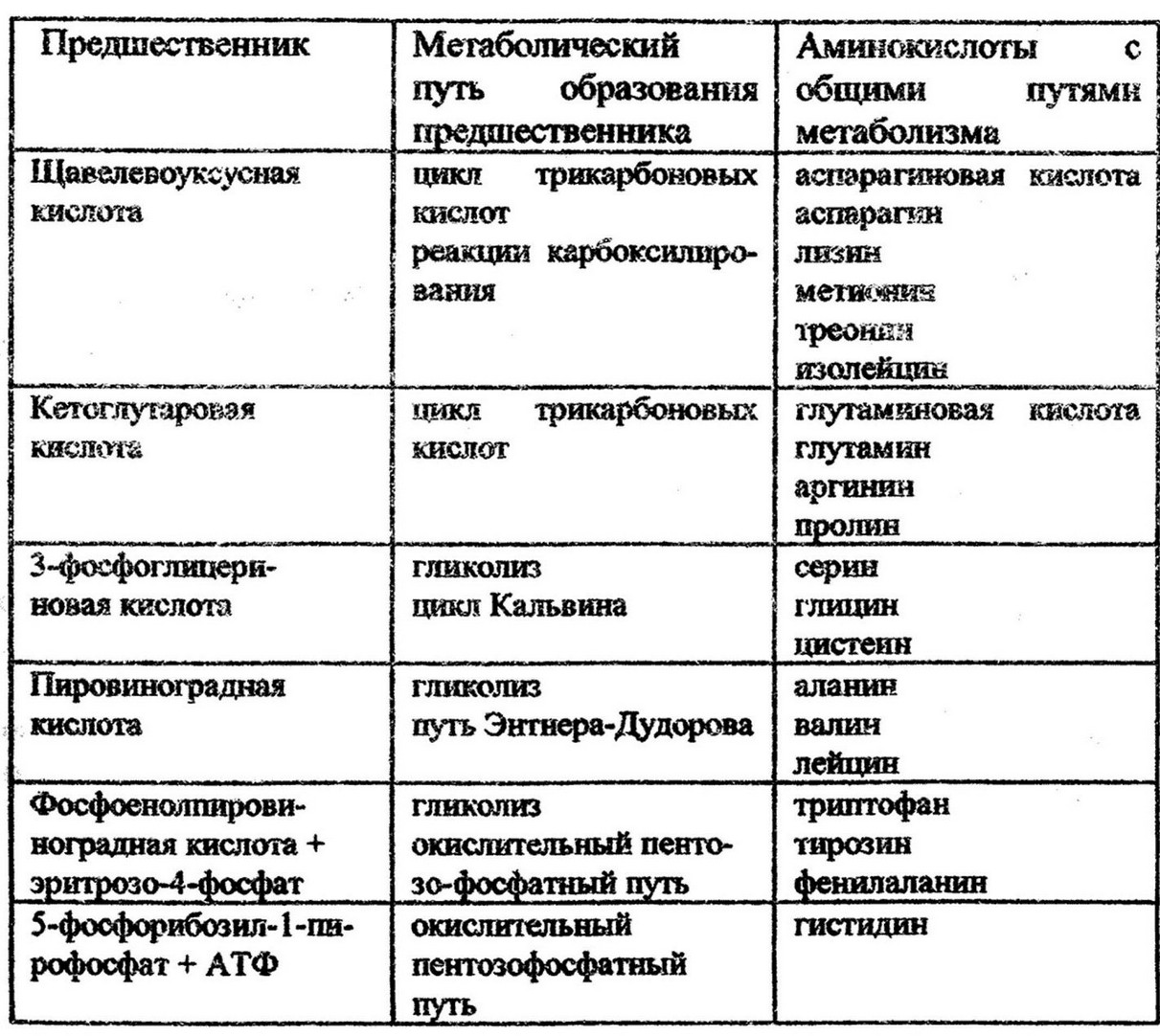
Особенностью биосинтеза аминокислот является наличие общих метаболических путей. Всего выделяют шесть таких путей и соответственно им все 20 аминокислот основного набора разделяют на шесть биосинтетических семейств (табл.З). Из них только гистин имеет самостоятельный путь биосинтеза.
Пути биосинтеза аминокислот приводят также к образованию иных азотсодержащих соединений, таких как полиамины, пурины, фолиевая кислота и другие вещества, необходимые для клетки.
Таблица 3
Особенности биосинтеза аминокислот
- Биосинтез белков
Аминокислоты, синтезированные внугриклеточно или потребленные из внешней среды, полимеризуются в молекулу белка. Механизм полимеризации довольно сложный и осуществляется в несколько этапов при непосредственном участии всех видов нуклеиновых кислот, которые играют решающую роль в биосинтезе белка. В ДНК закодирована информация о структуре каждого типа белка, характерного для определенного организма. Различные участки ДНК функционально неоднородны, и одна ее молекула может определять синтез большого числа функционально и химически различных белков клетки. Участок ДНК (несколько пар нуклеотидов), определяющий синтез одного типа белка, обозначается как ген. Ген передает информацию особой РНК3 которая синтезируется на нем как на матрице и называется информационной, или матричной, РНК (мРНК). При синтезе мРНК копируется нуклеотидная последовательность информации, закодированной ь ДНК, называемая транскрипцией. Процесс перевода нуклеотидной последовательности мРНК в последовательность аминокислот в полипептиде называется трансляцией. Таким образом, синтез полипептида включает два процесса – транскрипцию и трансляцию:
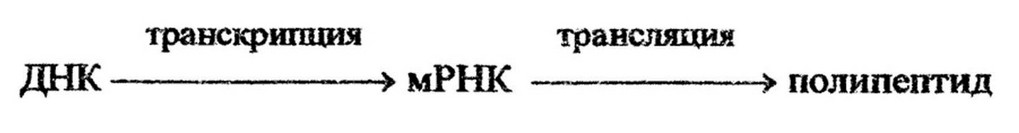
Включение конкретной аминокислоты в синтезируемую пептидную цепь определяется комбинацией трехсоседних нуклеотидов в молекуле мРНК – триплетом.
Триплеты ДНК называются кодонами. Большинство аминокислот кодируется несколькими триплетами. Это значит, что генетический код вырожден (табл.41
Генетический код
Таблица 4
| Первая | Вторая буква 1 | |||
| буква | У | ц | А | г ! |
| У | УУУФен1 | УЦУСер | УАУТир | УГУ Цис |
| УУЦФен | УЦЦСер | УАЦТир | УГЦЦис | |
| УУА Лей | УЦА Сер | УАА (нет)2 | УГА (нет)2 | |
| УУГ Лей | УЦГСер | УАГ (нет)2 | УПГТрн | |
| Ц | ЦУУ Лей | ПДУ Про | ЦАУ Гис | ЦГ У Apr |
| ЦУЦ Лей | ЦЦЦПро | ЦАЦ Гис | ЦГЦАрг | |
| ЦУА Дей | ЦЦАПро | ЦААГли | ЦТ A Apr | |
| ЦУГ Лей | ЦЦГПро | ЦАГГли | ЦГТ Apr | |
| А | АУУ Иле | АЦУТре | ААУ Асв | АГУ Сер |
| АУЦ Иле | АЦЦТре | ААЦ Асн | АЩСер | |
| АУА Иле | АДА Тре | АААЛиз | АГА Apr | |
| ДУГ Мег | АЦГТре | ААГ Лиз | ААГ Apr | |
| .. Г | ГУ У Вал | ЩУ Ала | ГАУ Асп | ГГУГлн 1 |
| ГУЦ Вал | ГЦЦАда | ГАД Асп | ГТЦ Гт | |
| 1 | ГУАЬал | ГИД Ала | ГААГлу | ГГ А Г ли |
| ГУГ Вал | ГЦГ Ала | ГАГ Гду | ПТ Гли 1 | |

Все молекулы мРНК содержат сигналы начала и конца кодируемого ими белка. Матричная РНК связывается с малой субчастицей рибосомы, объединяя группу рибосом в полисомы.
Синтез белка у всех организмов происходит на рибосомах (рис.7.8). Доставку аминокислот к рибосомам осуществляют транспортные РНК (тРНК) Каждая аминокислота доставляется к рибосомам специфичной для нее тРНК.
Первым этапом синтеза белка является активирование
Рис. 7.8. Схема биосинтеза белка
аминокислот и образование аминоацил – тРНК. Этот процесс катализируется специфичными для каждой аминокислоты ферментами аминоацил – тРНК-сннтетазами при участии АТФ в соответствии со следующим уравнением:
Транспортные РНК переносят аминокислоты к рибосомам, где они пептидными связями соединяются в пептидную цепь.
Перенос аминокислот к рибосомам посредством тРНК необходим, потому что сами по себе аминокислоты не способны узнавать кодоны в мРНК. Их узнают тРНК благодаря наличию в составе своей молекулы антикодонов. Кроме того, присоединение аминокислоты к рибозному остатку тРНК активирует карбоксильную группу аминокислоты и она способна к образованию пептидной связи. Узнавание кодона антикодоном (тРНК) контролируется рибосомами. Рибосомы движутся вдоль мРНК в направлениисчитывая кодоны путем
присоединения к ним соответствующей аминоацил-тРНК. С каждым присоединением аминокислоты рибосома передвигается на один триплет вдоль мРНК, «считывая» шаг за шагом всю информацию.
Когда рибосома доходит до последнего триплета, то после присоединения соответствующей ему аминокислоты к растущей полипеитидной цепи синтез белка прекращается.
Процесс трансляции, а следовательно, и образования полипегггидной цепи, очень сложен, В нем различают три стадии: инициацию, элонгацию и терминацию.
Инициация синтеза всех белков начинается со связывания N -фор мил “Метионил – тРНК с пептидильным центром (П) 30S-cy6- частицы рибосомы, которая присоединена к инициирующему триплету мРКК (АУГ или ГУГ). Затем нужная аминоamp;цил-тРНК подходит к аминоацильному центру (А) 50S-субчастицы рибосомы и образуется первая пептидная связь, Таким образом, стадия нжщиации включает все реакции, обеспечивающие формирование пептидной связи между первыми двумя аминокислотами.
Далее следует стадия элонгации. Ока включает реакции связывания соответствующих аминоацил-тРНК, образование пептидных связей и транслокацию. Пептидная цепь растет в направлении от Н-концевой аминокислоты к С-концевой, т. е. от аминокислоты со свободней аминогруппы к аминокислоте со свободной карбоксильной группой.
На стадии терминации полностью синтезированный полипептид освобождаете® от концевой тРНК, Рибосомы отделяются от мРНК к диссоциируют на исходные субчастицы gt; Терминацию синтеза белка осуществляют так называемые факторы освобождена. Это белки (RFi-‘l, RF-2), способные узнавать терминирующие кодоны (УАГ, УДА, УГА). Связывание фактора осгзобождения с тер ми и ир ующн м кодоном активирует
пептидилтрансферазу и она гидролизует связь между полипептидом и тРНК.
На рибосомах синтезируется белок первичной структуры, под которой понимают последовательность аминокислот в пептидной цепи. После завершения синтеза полипептидная цепь свертывается, приобретая вторичную и третичную структуру, а а ряде случаев объединяется с* другими полипептидными цепями и в результате образуется белок четвертичной структуры. Синтез белка происходит с расходованием большого количества энергии. Только для
образования одной пептидной связи затрачивается количество энергии, эквивалентное 4 молекулам АТФ.
- Биосинтез липидов
Микроорганизмы способны осуществлять биосинтез как сложных, так и простых липидов, которые входят в состав клеточной стенки и цитоплазматической мембраны, являются запасными веществами, компонентами пигментных систем и цепей электронного транспорта.
Источником для синтеза липидов микроорганизмами служат углеводы, спирты, органические кислоты. Кроме них, необходимо наличие в среде фосфатов для образования фосфор ил ированных предшественников биосинтеза.
Образованию липидов предшествует синтез эфиров жирных кислот и коэнзима А. Затем следуют реакции конденсации жирных кислот с глицерином, приводящие к появлению липидов. Пути синтеза липидов сложные, реакции катализируются многими ферментами и протекают с затратой значительных количеств энергии. Например, на синтез только стеариновой кислоты из глюкозы затрачивается 945,7 кал.
Глюкоза может служить также источником глицерина:
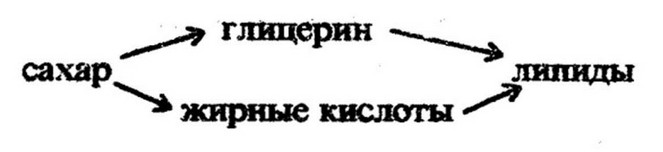
Глицерин получается в процессе анаэробного расщепления углеводов путем восстановления глицеринового альдегида. Жирные кислоты образуются из промежуточных продуктов распада углеводов, главным образом уксусной кислоты, уксусного альдегида, пировиноградной кислоты и этилового спирта. Важнейшей из них является уксусная кислота:
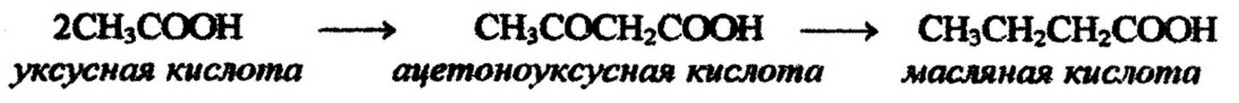
Но не сама уксусная кислота участвует в биосинтезе липидов, а ее производное – ацетилкоэнзим А, который является источником ацетильных радикалов и занимает центральное положение в метаболизме липидов:
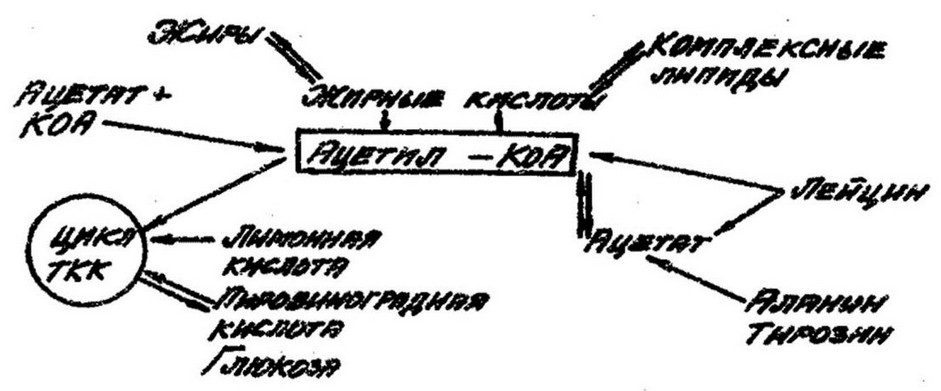
Макроэргическая тиоэфирная связь а цетил-К оА при
гидролизе высвобождает S200 кал. Данная энергия используется для переноса ацетильных остатков, необходимых для биосинтеза жирных кислот. Ацетилкоэнзим А за счет различных органических кислот при помощи ряда циклических процессов – конденсирования, гидрогенизации и дегидратации – обеспечивает образование длинных цепей жирных кислот, являющиеся компонентами лшшдов в клетках эубактерий.
У цианобактерий выявлены пали ненасыщенные жирные кислоты, имеющие две и более двойных связей, для образования которых аэробным бактериям необходим молекулярный кислород.
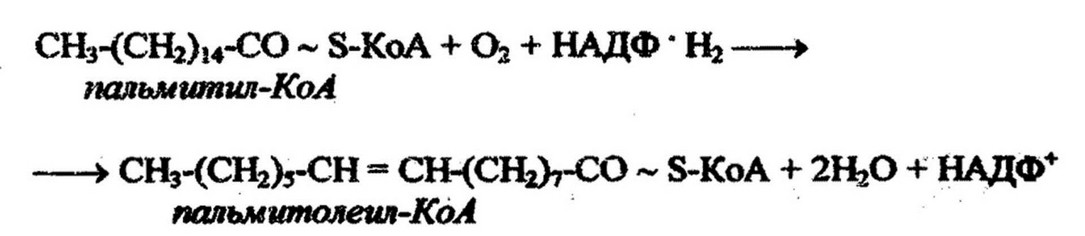
Второй путь введения двойной связи в молекулу кислоты при ее синтезе осуществляется у облигатно анаэробных и некоторых аэробных эубактерий в результате реакции дегидратации.
Синтез фосфолипидов состоит из нескольких этапов. Исходный субстрат фосфодиоксиацетон (промежуточный продукт гликолитического пути) восстанавливается до 3-фосфоглицерина к
которому присоединяются два остатка жирных кислот с образованием фосфатидной кислоты. Происходит активирование ее с помощью ЦТФ и присоединение к фосфатной группе серина, инозита, глицерина приводят к синтезу фосфатидил серина, фосфатидилинозита и фосфатидилглицерина.
Источник