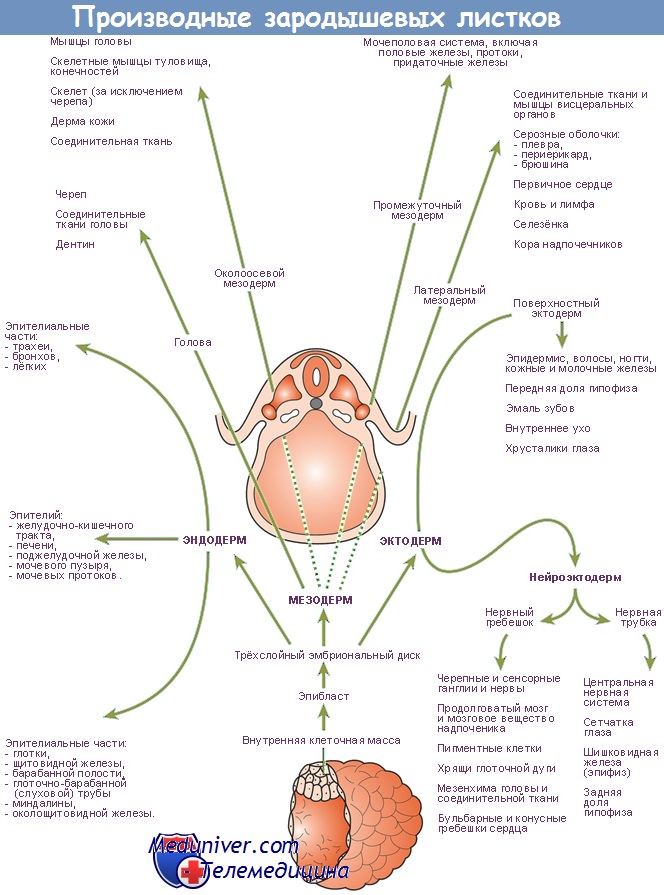
Кишечная трубка развивается из энтодермы

Развитие кишечной трубки. Формирование пищевода
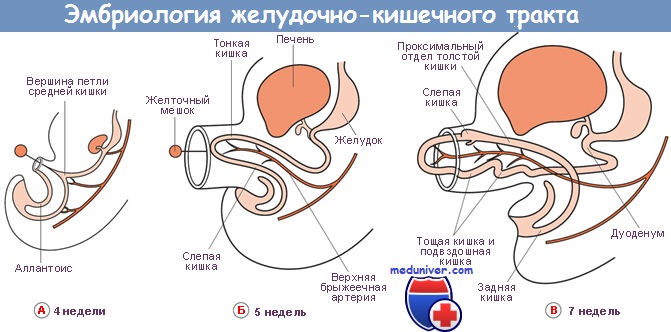
Если смотреть на продольный разрез человеческого эмбриона в возрасте приблизительно четырех недель и на модель его желудочно-кишечного тракта, то видно, что в данной стадии развития первичный кишечник имеет в общем прямую конфигурацию за исключением умеренного искривления, соответствующего искривлению всего тела, а также изгиба, в котором каудальный конец кишечной трубки загибается в хвостовой конец зародыша.
В диаметре просвета кишечной трубки еще не наблюдается значительных отклонений, за исключением веретенообразного расширения, представляющего закладку желудка, и менее значительного расширения в области задней кишки, называемого клоакой. Средний отдел кишки еще сообщается через сравнительно широкое отверстие с энтодермой желточного мешка.
На последующих стадиях развития происходят сложные изменения, касающиеся как самой кишечной стенки, так и ширины просвета и хода кишечной трубки. В результате этих процессов формируются отдельные части переваривающего отдела пищеварительной трубки и большой железы брюшной полости. Из энтодермы первичной кишечной трубки развиваются все эпителиальные и железистые элементы желудочно-кишечного тракта, а остальные компоненты его стенки (мускулатура, соединительная ткань) образуются из окружающей мезенхимы.
Из мезенхимной ткани состоят и первичные дорсальная и вентральная брыжейки (мезентерий) кишечной трубки, которые в виде двойной складки подвешивают ее к дорсальной и вентральной стенкам тела.
Формирование пищевода
Прямым продолжением каудальной части фарингеальной кишки является трубковидная закладка пищевода. В краниальной области от вентральной стенки данной закладки отщепляется зачаток дыхательных органов, развитие которых будет описано ниже. В каудальном направлении трубка будущего пищевода переходит в веретенообразное, мешковидное расширение будущего желудка.
Пищеводная трубка растет в длину, однако по отношению к более каудально расположенному отделу кишечной трубки она удлиняется незначительно. Приблизительно на шестой неделе развития стенка пищевода становится двуслойной. Его закладка располагается в широкой полосе мезенхимы висцерального мезобласта в области будущего средостения.
Двуслойный эпителий пищевода впоследствии превращается в типичный складчатый плоский эпителий. Пищеводные железы закладываются, начиная приблизительно с четвертого месяца, а первоначально круглый просвет пищевода, в связи с образованием слизистых и субмукозных складок, приобретает звездчатый на поперечном разрезе вид. Соединительнотканные и мышечные элементы пищеводной стенки, так же как и остальных отделов кишечной трубки, развиваются из окружающей мезенхимы.
Вместе с желудком, который в процессе своего развития, как это будет описано ниже, совершает поворот таким образом, что его дорсальная поверхность оказывается обращенной налево, а вентральная поверхность — направо. Этим поворотом объясняется и асимметричный ход обоих блуждающих нервов в данной области.
Учебное видео по развитию желудочно-кишечного тракта (эмбриогенезу)

– Вернуться в оглавление раздела “Акушерство.”
Оглавление темы “Формирование органов желудочно-кишечного тракта у эмбриона”:
1. Патология развития жаберных дуг. Развитие первичной полости рта
2. Развитие зубов. Этапы формирования зубов у эмбриона
3. Формирование коронки зуба. Развитие корней зубов
4. Формирование периодонта зуба. Развитие слюнных желез плода
5. Особенности развития крупных слюнных желез. Развитие области фарингеального кишечника
6. Фарингеальная кишка. Развитие языка эмбриона
7. Задняя закладка языка. Развитие внутренних жаберных борозд – мешков
8. Формирование cavum tympani и tuba pharyngotympanica. Развитие tonsilla palatum, thymus
9. Развитие околощитовидной и щитовидной желез. Формирование глотки
10. Развитие кишечной трубки. Формирование пищевода
Источник
Регуляция специализации энтодермы при формировании желудочно-кишечного тракта у плодаРазвитие ЖКТ происходит в три этапа. Первый этап — первичная специализация энтодермы. Второй этап заключается в формировании и структурировании кишечной трубки, что определяет передне-заднюю ось эмбриона и границы между различными органами. На третьем этапе начинается образование органов, которые в процессе развития выходят за границы кишечной трубки, например печени и поджелудочной железы. В экспериментах на лабораторных животных установлены эволюционно консервативные группы генов, принимающих участие в специализации энтодермы, и ряд других генов, участвующих только в процессе развития ЖКТ позвоночных. Эпителий этого тракта формируется из энтодермы — одного из эмбриональных зародышевых листков, возникающих в процессе гаструляции. Исследования, проведенные на таких лабораторных организмах, как круглые черви (почвенные нематоды Caenorhabdilis elegans), плодовые мушки (род Drosophila), африканские шпорцевые лягушки (род Xenopus), аквариумные рыбки Danio rerio и лабораторные мыши, позволили установить некоторые основные молекулярные механизмы регуляции процесса развития энтодермы. Моделирование на лабораторных мышах, у которых был проведен нокаут определенных генов для исследования их функций, в настоящее время рассматривается в качестве основы для изучения таких заболеваний ЖКТ у человека, как стеноз привратника, различные атрезии, неперфорированный анус. Специализация энтодермы может быть прослежена на самых ранних стадиях развития эмбриона. В классических экспериментальных исследованиях продемонстрирована способность эксплантатов куриных эмбрионов еще до процесса гаструляции, в культуре, к развитию ЖКТ, что свидетельствует о четкой детерминации процесса уже на данном этапе эмбрионального развития. Энтодерма куриных эмбрионов, выделенная и помещенная в лабораторных условиях в культуру до формирования ЖКТ, характеризуется экспрессией молекулярных маркеров, которые должны в норме появиться in vivo, что свидетельствует о наличии феномена ранней специализации. Появляются доказательства в поддержку гипотезы, что отличительные особенности энтодермы обусловлены функциональной клеточной независимостью, однако для нормального развития органов необходимо тесное взаимодействие энтодермы и мезодермы. В настоящее время на многочисленных лабораторных организмах установлены целые группы генов, способствующих специализации энтодермы. Один класс генов кодирует факторы транскрипции, которые, в свою очередь, непосредственно активируют гены-мишени. Второй класс генов кодирует синтез сигнальных молекул, обеспечивающих процесс межклеточной кооперации. Как минимум несколько факторов транскрипции, участвующих в процессе специализации энтодермы, продолжают экспрессироваться в тканях ЖКТ в течение всего последующего его развития, например семейства генов Fox и факторов GATA (семейство факторов транскрипции, характеризующихся способностью связываться с фрагментом дезоксирибонуклеиновой кислоты (ДНК) и с GATA-последовательностью нуклеотидов). В регуляции развития ЖКТ принимают участие, действуя в строго определенное время в различных локусах, сигнальные пути, или механизмы передачи регуляторных стимулов внутри клетки через определенную цепь молекул-посредников, представители группы факторов роста — трансформирующие факторы роста b в том числе непосредственно трансформирующий фактор роста b (TGF-b), и белок морфогенеза костной ткани (BMP), а также сигнальные пути hedgehog (сигнальные системы млекопитающих, представленные тремя различными белками). Продемонстрировано, что фактор транскрипции Sox-17 играет крайне важную роль в развитии энтодермы у мышей на ранних этапах эмбриогенеза, однако роль Sox-17 в дальнейшем не установлена. В последнее время выявлено, что сигнальный путь Notch (рецепторзависимый латентный фактор транскрипции, локализованный в клеточной цитоплазме) также выполняет важную функцию в регуляции развития ЖКТ. До настоящего времени не удалось выяснить, существует ли «главный ген», запускающий формирование энтодермы и последующее развитие ЖКТ. При исследовании некоторых животных моделей установлены гены, присутствие которых является необходимым и в то же время достаточным условием для формирования энтодермы, например ген mixer у шпорцевых лягушек. Ген мышей mixl1 играет основную роль в дифференцировке эмбриональных клеток в энтодерму. У других лабораторных животных обнаружены гены, необходимые для обеспечения данного процесса, однако недостаточные для формирования полноценной энтодермы. В исследованиях на лабораторных животных установлены начальные этапы индукции образования энтодермы. Энтодерма у мышей формируется из передней части первичной полоски эпибласта. Ключевой регулятор ранних этапов развития энтодермы — nodal-лиганд факторов роста суперсемейства TGF-b. Nodal экспрессируется в расположенном в бластодерме организационном центре или в узелке первичной бороздки эмбриона мыши. В отсутствие экспрессии nodal эмбрионы мыши не способны к гаструляции или к формированию дифинитивной энтодермы. Аналогично у эмбрионов Danio rerio в отсутствие экспрессии гомологов nodal также не образуется энтодерма. Напротив, экспрессия функционально активных рецепторов к TGF-b способствует преобразованию эмбриональных клеток в энтодерму. Реализация сигнального пути TGF осуществляется посредством стабильного комплекса промежуточных белков Smad. Необходимым для развития энтодермы является белок Smad2, и нокаут этого белка приводит к нарушению формирования энтодермы задней кишки, а разные мутации в гене, кодирующем Smad2, вызывают различные аномалии передней/головной кишки. FoxA2 также играет важную роль в развитии энтодермы, являясь мишенью сигнального пути, запущенного фактором nodal через белок Smad2. Smad2 и Smad3 совместно регулируют процесс развития энтодермы у мышей, оказывая различное влияние на формирование передней кишки и задней кишки. Кроме того, сигнальная система Smad необходима для развития печени. Основную роль в формировании печени играет ген Hex, экспрессия которого отсутствует либо существенно снижена при нарушенной продукции белков Smad. Этапы развития желудочно-кишечного тракта у плода
Установлено, что энтодерма находится в тесном контакте с мезодермой на протяжении всего ЖКТ. В экспериментальных исследованиях, выполненных на культурах тканей, было показано, что формирование энтодермы и ее дифференцировка в отдельные органы представляет собой результат взаимодействия мезодермы и энтодермы. Раньше всего было показано, что для формирования переднезадней оси энтодермы мышей необходимо сигнальное воздействие мезодермы на энтодерму посредством фактора роста фибробластов (FGF). Другие члены семейства FGF и рецепторы к ним необходимы для формирования печени. Еще три семейства генов крайне необходимы в процессе взаимодействия мезодермы и энтодермы — гены, кодирующие факторы сигнальных путей hedgehog и BMP, гены box. У почвенной нематоды Caenorhabditis elegans для специализации клеток, из которых развивается кишечный эпителий, необходимы два гена, кодирующие факторы транскрипции GATA, в то время как фактор GATA у плодовых мушек рода Drosophila кодируется одним геном serpent, наличие которого нужно для дифференцировки кишечной энтодермы. В кишечнике позвоночных экспрессируются три фактора группы GATA. Предполагается, что в процессах пролиферации и дифференцировки кишечного эпителия факторы GATA4, -5 и -6 выполняют различные функции, однако их функциональное значение на ранних этапах развития кишечной трубки у млекопитающих пока не установлено. Помимо факторов GATA, представители семейства forkhead-related факторов (кодируемые генами Fox) и сигнального пути WNT/Tcf являются важными регуляторами развития энтодермы. У позвоночных важную роль в инициации формирования энтодермы играют представители суперсемейства TGF-b. Молекула цитоскелета ELF3 задействована в сигнальном пути через факторы TGF-P, а также необходима для развития энтодермы. В отсутствие гена, кодирующего ELF3, у мышей полностью отсутствует кишечная энтодерма. Многочисленные факторы транскрипции, первоначально считавшиеся специфическими для формирования печени, играют роль и в развитии кишечника. При изучении эмбрионов мыши было отмечено, что некоторые из факторов транскрипции имеют характерные паттерны экспрессии, что позволило предположить их участие в развитии кишечника. Например, нуклеарный печеночный фактор (HNF) 3b (сейчас известный как FoxA2) оказался важным элементом в процессе самых ранних этапов дифференцировки ЖКТ и продолжает экспрессироваться в производных энтодермы у взрослых. Гомозиготные мутанты с полным отсутствием HNF3b не способны к формированию первичной полоски, предшествующей образованию кишечной трубки и ряда других структур. HNF3b участвует в образовании передней и средней, но не задней кишки. Идентифицировано большое количество факторов семейства Fox, некоторые из которых характеризуются преимущественной или специфической экспрессией в кишечнике. Один из представителей данного семейства, Foxl1, в норме экспрессируется кишечной мезодермой и представляет собой основной медиатор взаимодействия между эпителием и мезенхимой. Отсутствие указанного фактора сопровождается патологической пролиферацией эпителиальных клеток и нарушением развития кишечника. Этот фактор действует через экспрессию протеогликанов, проявляющих себя как корецепторы к компонентам сигнального пути WNT, и таким образом активирует комплекс WNT/P-катенин, регулирующий пролиферацию клеток. Группа исследователей во главе с Zaret предложила модель, в которой факторы FoxA2 и GATA открывают ДНК в клетках энтодермы, обеспечивая ее доступность для последующего связывания с факторами транскрипции, регулирующими клеточно-специфические гены. Таким образом, вероятно, при развитии кишечника многочисленные представители семейства факторов Fox играют важную роль в этом сложном процессе, который окончательно не изучен. У мышей некоторые гены, гомологичные гену caudal у дрозофил, экспрессируются исключительно в кишечнике. Cdx-1, экспрессирующийся в кишечнике взрослых организмов, также широко экспрессируется в тканях развивающегося эмбриона. Другой ген, Cdx-2, экспрессируется в висцеральной энтодерме на ранних стадиях эмбрионального развития, но впоследствии его паттерн экспрессии ограничен исключительно кишечником. Чрезмерная экспрессия Cdx-2 приводит к дальнейшей дифференцировке популяции клеток кишечника, которые в норме должны оставаться недифференцированными. В то же время эктопическая экспрессия Cdx-2 в стенке желудка приводит к формированию ткани, аналогичной кишечнику. Cdx-2 представляет собой важный фактор дифференцировки кишечника, однако до настоящего времени его роль в формировании кишечной трубки на ранних стадиях эмбрионального развития точно не установлена.
– Также рекомендуем “Генетическая регуляция формирования кишечной трубки” Оглавление темы “Эмбриогенез желудочно-кишечного тракта”:
|
Источник
Генетическая регуляция формирования кишечной трубкиКишечная трубка формируется из слоя энтодермы за счет ее сворачивания, которое начинается при образовании туловищных складок кишечных ворот на переднем и заднем концах эмбриона. Взаимодействие между энтодермой и мезодермой играет решающую роль в процессе развития ЖКТ. Ключевой механизм, выступающий в качестве медиатора взаимодействия энтодермы и мезодермы в развитии ЖКТ, — система сигнальных белков Sonic hedgehog (Shh) и Indian hedgehog (Ihh). Как Shh, так и Ihh играют важную роль в структурировании развивающегося ЖКТ по переднезадней оси и в радиальном направлении и влияют на развитие мышечной ткани из мезодермы. Одна из мишеней сигнального пути hegdehog — группа сигнальных молекул BMP. Shh сначала экспрессируется в примитивной энтодерме эмбриона, затем в энтодерме конечных отделов передней и задней кишки, а впоследствии — во всей кишечной энтодерме, а также в области крипт у взрослых организмов. ВМР-4 экспрессируется в мезодерме, прилежащей к конечным отделам передней и задней кишки, однако возможна и его эктопическая экспрессия в висцеральной мезодерме, индуцируемая белком Shh. Энтодерма конечных отделов кишки синтезирует белок Shh. Таким образом, Shh играет главную регулирующую роль в развитии как передней, так и задней кишки. При отсутствии продукции данного белка у мышей аномально развивается передняя кишка, например происходит атрезия пищевода и возникают трахеопищеводные свищи, появляются аномалии задней кишки, в частности персистирующая клоака. Более того, необходимо присутствие факторов транскрипции Gli2 и Gli3, выполняющих передаточную функцию для сигнального белка Shh. У мышей при нокауте фактора транскрипции Gli2 и, соответственно, сниженной экспрессии Gli3 наблюдают атрезию пищевода и формирование трахеопищеводных свищей. Установлено отсутствие перфорирования ануса в сочетании с прямокишечно-уретральным свищом и анальным стенозом у мышей при отсутствии экспрессии Gli2 либо Gli3. Роль данного сигнального пути в развитии желудочно-кишечного тракта у мышей предположительно ограничена первой половиной срока гестации, т.к. в эксперименте введение через 12,5 сут эмбрионального развития блокирующих антител к белкам hedgehog не оказывало влияния на морфологию кишки. Процессы пролиферации в области крипт и метаболизма липидов оказались нарушенными, что свидетельствует о значимости белков hedgehog для более поздних этапов развития организма.
Учебное видео по развитию желудочно-кишечного тракта (эмбриогенезу)
– Также рекомендуем “Генетическая регуляция формирования органов желудочно-кишечного тракта” Оглавление темы “Эмбриогенез желудочно-кишечного тракта”:
|
Источник