Кишечный эпителий из энтодермы

Энтодерма — внутренний слой гас-трулы (внутренний зародышевый листок).[ …]
Энтодерма образует внутренний эпителиальный покров кишечника и желез системы его, развившихся путем выпячивания участков кишечника, а равно и плавательного пузыря. Из мезодермы развивается сперва хорда, около нее позвонки, все кости скелета, чешуя и кожные жучки рыб, гладкие и поперечнополосатые мышцы, эпителий полости тела, опорная ткань органов (соединительная ткань), наружная стенка кишечника, сосудистая и лимфатическая система с кровью и лимфой, половые клетки (яйцевые и семенные), почки.[ …]
Мезодерма — слой зародышевых клеток между эктодермой и энтодермой.[ …]
У кишечнополостных тело состоит из двух слоев — эктодермы и энтодермы, представляющих собой наружный и внутренний эпителиальные слои. Наружные эпителиальные клетки являются стрекательными клетками, содержащими ядовитую жидкость, тогда как внутренние эпителиальные клетки секретируют пищеварительные ферменты и обеспечивают пищеварение. Поэтому предполагают, что первыми сформировались слои эпителиальных клеток и их роль в эволюции многоклеточных аналогизируется с ролью клеточных стенок и мембран одноклеточных организмов.[ …]
Образование тканей (гистогенез) у животных происходит из эктодермы, энтодермы, мезодермы и мезенхимы в период эмбриогенеза, а основными элементами тканей, как отмечено выше, являются клетки и их производные в виде неклеточных структур. Таким образом, ткань можно определить в виде сообщества клеток и их производных со специализированными функциями.[ …]
Но во всех случаях гаструла существует и состоит из двух слоев клеток, составляющих эктодерму и энтодерму; последняя ограничивает полость, первичный кишечник гаструлы, открывающуюся наружу первичным ртом — бластопорусом. Полость гаструлы наполнена питательным желтком, образующим в бласто-пору се „желточную пробку“. По мере роста эктодермы и энтодермы бла-стопор замыкается, и тогда весь питательный желток находится в за мкнутой полости желудка зародыша; у костистых рыб с большим количеством питательного желтка он еще долгое время существует у малька в виде так называемого „желточного пузыря“.[ …]
Начало изучению «зачатковых» или первичных половых клеток (ППК) у рыб было положено М. Нуссбаумом, который описал возникновение их у эмбрионов форели в области энтодермы и миграцию к месту формирования гонад (Nussbaum, 1880). Он же был первым исследователем, применившим идею раннего обособления ППК и экстрагонадного их происхождения к позвоночным животным.[ …]
Ко времени зарастания бласто-пора или раньше того начинается очень важный процесс, именно образование третьего промежуточного эмбрионального пласта—мезодермы, врастающего в щель между экто -и энтодермой.[ …]
Развитие двух зародышевых листков характерно для губок и кишечнополостных. Однако для хордовых в период гаструляции характерно развитие третьего зародышевого листка — мезодермы (от греч. теэоз — средний), образующегося между эктодермой и энтодермой (рис. 90).[ …]
Тип Кишечнополостные (Сое1еп1ега1а). Организмы этого типа являются обитателями в основном морей, но они проникли и в пресные воды. Известно около 9000 видов. Кишечнополостным присуща довольно простая организация. Для них характерна радиально-осевая симметрия. Их тело состоит из экто- и энтодермы, между которыми находится мезоглея, представляющая собой слой неклеточного вещества.[ …]
Развитие (гаструляция) изолецитальных яиц происходит путем инвагинации (впячивания) вегетативного полюса внутрь бластулы, в результате чего противоположные полюса почти сливаются, а бластоцель (полость бластулы) почти исчезает либо полностью исчезает. Внешний слой клеток зародыша получил название эктодермы (от греч. ectos — снаружи, derma — кожа) или наружного зародышевого листка, тогда как внутренний — энтодермы (от греч. entos — внутри) или внутреннего зародышевого листка. Образующаяся при этом полость получила название гастроцели, или первичной кишки, вход в которую называют бластопором (первичным ртом).[ …]
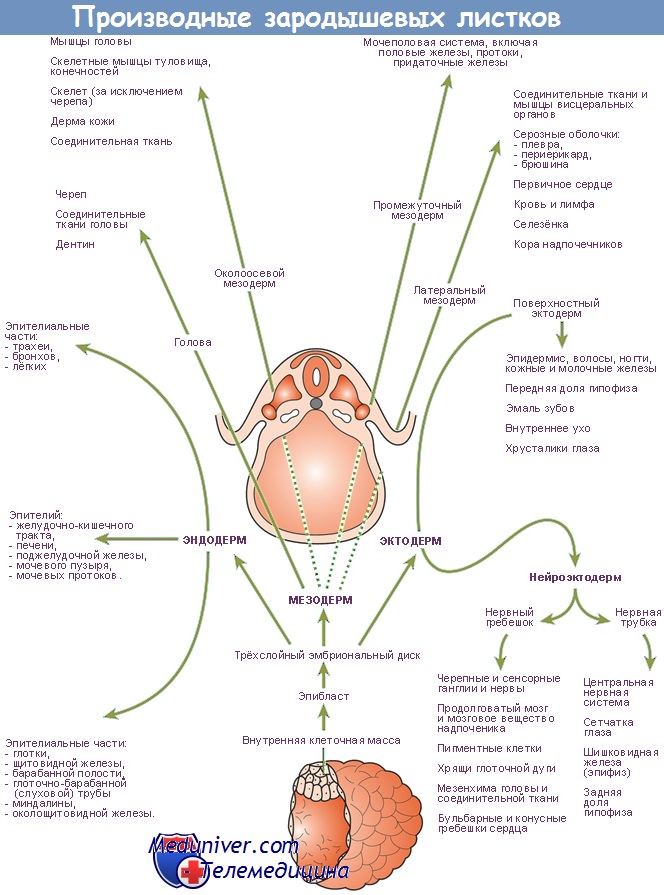
Развитие (дифференцировка) зародышевых листков в ходе эмбриогенеза сопровождается тем, что из них формируются различные ткани и органы. В частности, из эктодермы развиваются эпидермис кожи, ногти и волосы, сальные и потовые железы, нервная система (головной мозг, спинной мозг, ганглии, нервы), рецепторные клетки органов чувств, хрусталик глаза, эпителий рта, носовой полости и анального отверстия, зубная эмаль. Из энтодермы развиваются эпителий пищевода, желудка, кишек, желчного пузыря, трахеи, бронхов, легких, мочеиспускательного канала, а также печень, поджелудочная железа, щитовидная, паращитовидная и зобная железы. Из мезодермы развиваются гладкая мускулатура, скелетные и сердечные мышцы, дерма, соединительная ткань, кости й хрящи, дентин зубов, кровь и кровеносные сосуды, брыжейка, почки, семенники и яичники. У человека первыми обособляются головной и спинной мозг. Через 26 дней после овуляции длина человеческого зародыша составляет около 3,5 мм. При этом уже видны зачатки рук, но зачатки ног только вступают в развитие. Через 30 дней после овуляции длина зародыша равна уже 7,5 мм. В это время уже можно различить сегментацию зачатков конечностей, глазные бокалы, полушария головного мозга, печень, желчный пузырь и даже разделение сердца на камеры.[ …]
Эпителиальная ткань, или эпителий, состоит из клеток, покрывающих поверхность тела, внутренние поверхности внутренних органов (желудок, мочевой пузырь и др.), поверхности серозных оболочек (брюшина, плевра, перикард), а также из клеток, образующих некоторые железы (слюнные железы, поджелудочная железа и др.). Поэтому различают покровный и железистый (секреторный) эпителий. Из эктодермы развивается эпителий кожи, из энтодермы — эпителий желудка, кишечника, легких и др., а из мезодермы — эпителий почек, серозных оболочек и других структур.[ …]
Внешним признаком развития икринки является скопление плазмы на анимальном полюсе и образование бластодиска. Развитие идет по следующей схеме: дробление бластодиска (с образованием сначала крупноклеточной, затем мелкоклеточной морулы); появление бластулы, внутри которой имеется первичная полость тела — бластоцель; в результате продолжающегося размножения клеток — наступление гаструляции, в процессе которой клетки анимального полюса надвигаются на желток, образуются два зародышевых листка (экто- и энтодерма), полость гаструлы представляет собой первичную полость кишечника. Далее зародышевые листки дифференцируются на зачатки тканей и органов (рис. 3).[ …]
Данные эмбриологии свидетельствуют о том, что после гаструляции развитие зародышей проходит в одном из двух направлений. С одной стороны, бластопор превращается либо в анальное отверстие, либо занимает положение около этого отверстия, как это имеет место в случае иглокожих и хордовых. С другой стороны, бластопор развивается в ротовое отверстие, либо занимает положение около этого отверстия, как это имеет место в случае кольчатых червей, моллюсков, членистоногих и др. При развитии как в одном направлении, так и другом между эктодермой и энтодермой развивается мезодерма, которая у иглокожих и позвоночных закладывается частично (карманами или выпячиваниями первичной кишки), а у кольчатых червей и других организмов закладывается из специальных клеток, дифференцирующихся на ранних этапах развития.[ …]
После оплодотворения икринки набухают, в них происходит обводнение желтка с образованием вокруг него перивителлинового пространства, заполненного жидкостью. Перивителлиновое пространство образуется и в неоплодотворенных икринках, после чего они обычно погибают. Яйцеклетки костистых рыб относятся к те-лолецитальному типу. В них ядро и плазма клетки находятся на анимальном полюсе, а желток — на противоположной части (вегетативном полюсе). В результате дробление охватывает только верхнюю часть клетки, начинаясь с образования бластодиска. Дальше развитие идет по общей схеме: образование бластулы, морулы, гаст-рулы. При гаструляции клетки анимального полюса окружают жел-ток с образованием двух зародышевых листков (экто- и энтодермы). Далее развитие переходит в стадию органогенеза, подвижного эмбриона, заканчиваясь выклевом предличинок. Продолжительность инкубации икры у разных видов рыб различная и зависит от температуры воды. Например, при оптимальных температурах (20— 22 °С) икра карпа развивается в течение 3-4 сут, а радужной форели при температуре около 10 °С — в течение 45-60 сут.[ …]
Онтогенез. Эмбриональное развитие организма
Необходимо запомнить
ВАЖНО!
Онтогенез – индивидуальное развитие организма.
Периоды онтогенеза
Эмбриональный – от образования зиготы до рождения или выхода из яйцевых оболочек.
Постэмбриональный – от выхода из яйцевых оболочек или рождения, до смерти.
Эмбриональный период развития у животных:
Дробление начинается через несколько часов после образования зиготы, образовавшейся в результате оплодотворения. Первое деление происходит в вертикальной плоскости, и образуются две одинаковые клетки – бластомеры. Они не расходятся, а делятся ещё раз, в результате чего образуются уже 4 бластомера. Далее они делятся, но уже в горизонтальной плоскости. Деление бластомеров быстро следуют одно за другим, и они не успевают расти. Поэтому на начальных стадиях дробления комочек бластомеров, который называется морула, не превосходит по своим размерам зиготу. Образуется однослойный зародыш – бластула. Полость внутри бластулы – бластоцель.
Гаструляция
Стенки гаструлы состоят из двух слоёв клеток – эктодермы и энтодермы, называемых зародышевыми листками. Полость внутри гаструлы называется первичной кишкой, а отверстие, которое в неё ведёт – первичным ртом.
У вторичноротых животных, к которым относятся все хордовые, на месте первичного рта образуется анальное отверстие, а на противоположном полюсе зародыша – вторичный рот. На этой стадии появляются первые признаки дифференцировки клеток.
Нейрула – трёхслойный зародыш. Из клеток, лежащих между экто – и энтодермой у всех многоклеточных животных за исключением кишечнополостных и губок образуется третий зародышевый листок – мезодерма. На стадии нейрулы начинается первичный органогенез. Дальнейшая дифференцировка клеток зародыша приводит к возникновению многочисленных производных зародышевых листков – органов и тканей. Таким образом, уже на ранних стадиях эмбрионального развития онтогенеза из внешне одинаковых бластомеров развиваются различные по строению и функциям ткани, органы и системы органов. Этот процесс получил название дифференцировки клеток. При дальнейшей дифференцировке клеток, входящих в состав зародышевых листков, образуются органы.
В процессе эмбрионального развития ткани зародыша оказывают влияние друг на друга – эмбриональная индукция.
Производные эктодермы – наружный слой кожи – эпителий и его производные: волосы, ногти, когти, рога, копыта, чешуя рыб, пресмыкающихся, кожные железы, нервная система, эмаль зубов, производные кожи: органы чувств: глаза, уши и др.
Производные энтодермы – эпителий внутренних органов: кишечника, жабр, лёгких. Пищеварительные железы – печень, поджелудочная железа.
Производные мезодермы хрящевая и костная ткань, мышцы, почки, сердечно – сосудистая система, половые железы, дентин зубов.
На развивающийся зародыш оказывает влияние окружающая среда. В большей степени эта зависимость проявляется у беспозвоночных животных. У плацентарных млекопитающих посредником между зародышем и окружающей средой является организм матери, от которого эмбрион получает питание, кислород, тепло.
Основателем современной эмбриологии является российский учёный К. М. Бэр. В 1828 г. он опубликовал сочинение «История развития животных».
Заслуга создания эволюционной эмбриологии также принадлежит замечательным русским учёным А. О. Ковалевскому, И. И. Мечникову, А. Н. Северцову, И. И. Шмальгаузену. Современным представлениям о зародышевых листках, наука обязана А.О. Ковалевскому, который обнаружил эктодерму, энтодерму и мезодерму у всех групп хордовых.
Немецкие учёные Ф. Мюллер и Э. Геккель сформулировали биогенетический закон, согласно которому онтогенез, т. е. индивидуальное развитие вида, есть краткое повторение филогенеза – исторического развития вида которому он относится. В 1866 Геккель вводит понятие онтогенез.
Российский учёный – академик А. Н. Северцов установил, что в индивидуальном развитии животных повторяются признаки не взрослых предков, а их зародышей.
Галерея открытий
Зародышевые листки и их производные
Источник
Источник
Эпителии кишечного типа. Эпителии почечного типа.
В кишечном типе эпителиев эпителиальный дифферон развивается из материала кишечной энтодермы. Наиболее общий гистологический признак эпителиев кишечного типа — однослойность и высокопризматическая форма эпителиоциов. Вместе с тем каждая разновидность кишечных эпителиев имеет свои органоспецифические особенности строения, функции и гистотопографии. Примером эпителия данного типа является всасывающий эпителий слизистой оболочки тонкой кишки. Это однослойный цилиндрический эпителий с гетерополярностью — различным строением базальной и апикальной частей клеток. На апикальной поверхности клеток имеются микроворсинки, которые образуют щеточную каемку. При этом всасывающая поверхность увеличивается в 25-30 раз.
В надмембранном комплексе — гликокаликсе — располагаются ферменты пристеночного пищеварения. Для эпителиев, образующих выстилку пищеварительного канала, характерно сильное развитие плотных межклеточных контактов запирающего типа, благодаря чему эпителиальный слой выполняет барьерную функцию. Внутрь организма вещества поступают не по межклеточным пространствам, которые прочно перегорожены замыкательными поясками, а непосредственно через сами клетки эпителия.
В составе эпителия существует другая разновидность клеток — бокаловидные экзокриноциты — это слизистые внутриэпителиальные одноклеточные железы. В их цитоплазме содержится много слизистого секрета, ядро оттеснено в базальную часть.
Эпителий состоит из разных по степени созревания клеток: стволовых, камбиальных, малодифференцированных, дифференцированных (зрелых) и заканчивающих жизненный цикл. Стволовые клетки способны к дивергентной дифференцировке и формированию дифферонов каемчатых, апикально-зернистых эпителиоцитов, бокаловидных экзокриноцитов и эндокриноцитов.
В процессе физиологической регенерации эпителиальный пласт обновляется в течение 3-5 суток.
К эпителиям кишечного типа относятся также эпителиальные ткани, составляющие основную массу печени и поджелудочной железы. Эпителии этих органов развиваются в эмбриогенезе из общего с кишечным эпителием энтодермального зачатка и представляют особые разновидности эпителиев кишечного типа. В их структуре важный гистологический признак — расположение клеток в виде пласта — наблюдается лишь на ранних этапах гисто- и органогенеза. В процессе последующего гистогенеза их эпителии приобретают специфические для желез черты строения, расположения и функции.
Эпителии почечного типа
Почечный тип эпителиев развивается из мезодермы, точнее нефротомов — сегментных ножек. Эпителии почечного типа образуют стенку мочевых канальцев почки. По своему строению — это однослойные эпителии. По длине мочевых канальцев эпителий имеет разное строение. Эпителиоциты могут быть плоские, кубические, призматические и различаются по функции. Одни из них выполняют функцию фильтрации, другие — всасывающую, третьи — секреторную функцию. В зрелом организме митотическая активность эпителиоцитов крайне низкая.
– Также рекомендуем “Эпителии целомического типа. Эпителии нейроглиального типа.”
Оглавление темы “Виды тканей. Кровь.”:
1. Дифференциация клеток. Детерминация клеток. Клеточная гибель. Понятие о диффероне.
2. Классификация тканей. Виды тканей.
3. Эпителиальные ткани. Эпителий.
4. Эпителии кожного типа. Эпидермис. Многослойные эпителии.
5. Эпителии кишечного типа. Эпителии почечного типа.
6. Эпителии целомического типа. Эпителии нейроглиального типа.
7. Железистые эпителии. Физиологическая регенерация эпителиев. Ткани внутренней среды.
8. Мезенхима. Ретикулярная ткань.
9. Кровь. Плазма крови. Эритроциты человека.
10. Лейкоциты. Нейтрофилы. Эозинофильные гранулоциты.
Источник
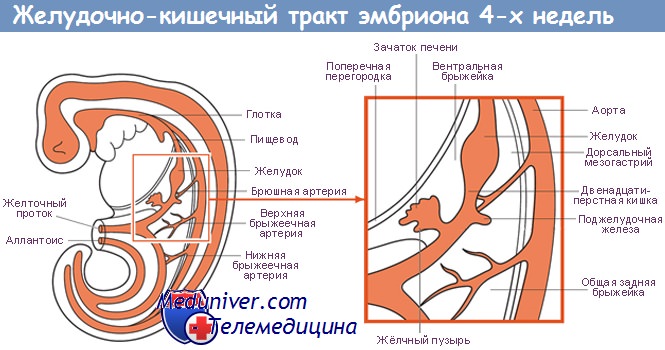
Регуляция специализации энтодермы при формировании желудочно-кишечного тракта у плодаРазвитие ЖКТ происходит в три этапа. Первый этап — первичная специализация энтодермы. Второй этап заключается в формировании и структурировании кишечной трубки, что определяет передне-заднюю ось эмбриона и границы между различными органами. На третьем этапе начинается образование органов, которые в процессе развития выходят за границы кишечной трубки, например печени и поджелудочной железы. В экспериментах на лабораторных животных установлены эволюционно консервативные группы генов, принимающих участие в специализации энтодермы, и ряд других генов, участвующих только в процессе развития ЖКТ позвоночных. Эпителий этого тракта формируется из энтодермы — одного из эмбриональных зародышевых листков, возникающих в процессе гаструляции. Исследования, проведенные на таких лабораторных организмах, как круглые черви (почвенные нематоды Caenorhabdilis elegans), плодовые мушки (род Drosophila), африканские шпорцевые лягушки (род Xenopus), аквариумные рыбки Danio rerio и лабораторные мыши, позволили установить некоторые основные молекулярные механизмы регуляции процесса развития энтодермы. Моделирование на лабораторных мышах, у которых был проведен нокаут определенных генов для исследования их функций, в настоящее время рассматривается в качестве основы для изучения таких заболеваний ЖКТ у человека, как стеноз привратника, различные атрезии, неперфорированный анус. Специализация энтодермы может быть прослежена на самых ранних стадиях развития эмбриона. В классических экспериментальных исследованиях продемонстрирована способность эксплантатов куриных эмбрионов еще до процесса гаструляции, в культуре, к развитию ЖКТ, что свидетельствует о четкой детерминации процесса уже на данном этапе эмбрионального развития. Энтодерма куриных эмбрионов, выделенная и помещенная в лабораторных условиях в культуру до формирования ЖКТ, характеризуется экспрессией молекулярных маркеров, которые должны в норме появиться in vivo, что свидетельствует о наличии феномена ранней специализации. Появляются доказательства в поддержку гипотезы, что отличительные особенности энтодермы обусловлены функциональной клеточной независимостью, однако для нормального развития органов необходимо тесное взаимодействие энтодермы и мезодермы. В настоящее время на многочисленных лабораторных организмах установлены целые группы генов, способствующих специализации энтодермы. Один класс генов кодирует факторы транскрипции, которые, в свою очередь, непосредственно активируют гены-мишени. Второй класс генов кодирует синтез сигнальных молекул, обеспечивающих процесс межклеточной кооперации. Как минимум несколько факторов транскрипции, участвующих в процессе специализации энтодермы, продолжают экспрессироваться в тканях ЖКТ в течение всего последующего его развития, например семейства генов Fox и факторов GATA (семейство факторов транскрипции, характеризующихся способностью связываться с фрагментом дезоксирибонуклеиновой кислоты (ДНК) и с GATA-последовательностью нуклеотидов). В регуляции развития ЖКТ принимают участие, действуя в строго определенное время в различных локусах, сигнальные пути, или механизмы передачи регуляторных стимулов внутри клетки через определенную цепь молекул-посредников, представители группы факторов роста — трансформирующие факторы роста b в том числе непосредственно трансформирующий фактор роста b (TGF-b), и белок морфогенеза костной ткани (BMP), а также сигнальные пути hedgehog (сигнальные системы млекопитающих, представленные тремя различными белками). Продемонстрировано, что фактор транскрипции Sox-17 играет крайне важную роль в развитии энтодермы у мышей на ранних этапах эмбриогенеза, однако роль Sox-17 в дальнейшем не установлена. В последнее время выявлено, что сигнальный путь Notch (рецепторзависимый латентный фактор транскрипции, локализованный в клеточной цитоплазме) также выполняет важную функцию в регуляции развития ЖКТ. До настоящего времени не удалось выяснить, существует ли «главный ген», запускающий формирование энтодермы и последующее развитие ЖКТ. При исследовании некоторых животных моделей установлены гены, присутствие которых является необходимым и в то же время достаточным условием для формирования энтодермы, например ген mixer у шпорцевых лягушек. Ген мышей mixl1 играет основную роль в дифференцировке эмбриональных клеток в энтодерму. У других лабораторных животных обнаружены гены, необходимые для обеспечения данного процесса, однако недостаточные для формирования полноценной энтодермы. В исследованиях на лабораторных животных установлены начальные этапы индукции образования энтодермы. Энтодерма у мышей формируется из передней части первичной полоски эпибласта. Ключевой регулятор ранних этапов развития энтодермы — nodal-лиганд факторов роста суперсемейства TGF-b. Nodal экспрессируется в расположенном в бластодерме организационном центре или в узелке первичной бороздки эмбриона мыши. В отсутствие экспрессии nodal эмбрионы мыши не способны к гаструляции или к формированию дифинитивной энтодермы. Аналогично у эмбрионов Danio rerio в отсутствие экспрессии гомологов nodal также не образуется энтодерма. Напротив, экспрессия функционально активных рецепторов к TGF-b способствует преобразованию эмбриональных клеток в энтодерму. Реализация сигнального пути TGF осуществляется посредством стабильного комплекса промежуточных белков Smad. Необходимым для развития энтодермы является белок Smad2, и нокаут этого белка приводит к нарушению формирования энтодермы задней кишки, а разные мутации в гене, кодирующем Smad2, вызывают различные аномалии передней/головной кишки. FoxA2 также играет важную роль в развитии энтодермы, являясь мишенью сигнального пути, запущенного фактором nodal через белок Smad2. Smad2 и Smad3 совместно регулируют процесс развития энтодермы у мышей, оказывая различное влияние на формирование передней кишки и задней кишки. Кроме того, сигнальная система Smad необходима для развития печени. Основную роль в формировании печени играет ген Hex, экспрессия которого отсутствует либо существенно снижена при нарушенной продукции белков Smad. Этапы развития желудочно-кишечного тракта у плода
Установлено, что энтодерма находится в тесном контакте с мезодермой на протяжении всего ЖКТ. В экспериментальных исследованиях, выполненных на культурах тканей, было показано, что формирование энтодермы и ее дифференцировка в отдельные органы представляет собой результат взаимодействия мезодермы и энтодермы. Раньше всего было показано, что для формирования переднезадней оси энтодермы мышей необходимо сигнальное воздействие мезодермы на энтодерму посредством фактора роста фибробластов (FGF). Другие члены семейства FGF и рецепторы к ним необходимы для формирования печени. Еще три семейства генов крайне необходимы в процессе взаимодействия мезодермы и энтодермы — гены, кодирующие факторы сигнальных путей hedgehog и BMP, гены box. У почвенной нематоды Caenorhabditis elegans для специализации клеток, из которых развивается кишечный эпителий, необходимы два гена, кодирующие факторы транскрипции GATA, в то время как фактор GATA у плодовых мушек рода Drosophila кодируется одним геном serpent, наличие которого нужно для дифференцировки кишечной энтодермы. В кишечнике позвоночных экспрессируются три фактора группы GATA. Предполагается, что в процессах пролиферации и дифференцировки кишечного эпителия факторы GATA4, -5 и -6 выполняют различные функции, однако их функциональное значение на ранних этапах развития кишечной трубки у млекопитающих пока не установлено. Помимо факторов GATA, представители семейства forkhead-related факторов (кодируемые генами Fox) и сигнального пути WNT/Tcf являются важными регуляторами развития энтодермы. У позвоночных важную роль в инициации формирования энтодермы играют представители суперсемейства TGF-b. Молекула цитоскелета ELF3 задействована в сигнальном пути через факторы TGF-P, а также необходима для развития энтодермы. В отсутствие гена, кодирующего ELF3, у мышей полностью отсутствует кишечная энтодерма. Многочисленные факторы транскрипции, первоначально считавшиеся специфическими для формирования печени, играют роль и в развитии кишечника. При изучении эмбрионов мыши было отмечено, что некоторые из факторов транскрипции имеют характерные паттерны экспрессии, что позволило предположить их участие в развитии кишечника. Например, нуклеарный печеночный фактор (HNF) 3b (сейчас известный как FoxA2) оказался важным элементом в процессе самых ранних этапов дифференцировки ЖКТ и продолжает экспрессироваться в производных энтодермы у взрослых. Гомозиготные мутанты с полным отсутствием HNF3b не способны к формированию первичной полоски, предшествующей образованию кишечной трубки и ряда других структур. HNF3b участвует в образовании передней и средней, но не задней кишки. Идентифицировано большое количество факторов семейства Fox, некоторые из которых характеризуются преимущественной или специфической экспрессией в кишечнике. Один из представителей данного семейства, Foxl1, в норме экспрессируется кишечной мезодермой и представляет собой основной медиатор взаимодействия между эпителием и мезенхимой. Отсутствие указанного фактора сопровождается патологической пролиферацией эпителиальных клеток и нарушением развития кишечника. Этот фактор действует через экспрессию протеогликанов, проявляющих себя как корецепторы к компонентам сигнального пути WNT, и таким образом активирует комплекс WNT/P-катенин, регулирующий пролиферацию клеток. Группа исследователей во главе с Zaret предложила модель, в которой факторы FoxA2 и GATA открывают ДНК в клетках энтодермы, обеспечивая ее доступность для последующего связывания с факторами транскрипции, регулирующими клеточно-специфические гены. Таким образом, вероятно, при развитии кишечника многочисленные представители семейства факторов Fox играют важную роль в этом сложном процессе, который окончательно не изучен. У мышей некоторые гены, гомологичные гену caudal у дрозофил, экспрессируются исключительно в кишечнике. Cdx-1, экспрессирующийся в кишечнике взрослых организмов, также широко экспрессируется в тканях развивающегося эмбриона. Другой ген, Cdx-2, экспрессируется в висцеральной энтодерме на ранних стадиях эмбрионального развития, но впоследствии его паттерн экспрессии ограничен исключительно кишечником. Чрезмерная экспрессия Cdx-2 приводит к дальнейшей дифференцировке популяции клеток кишечника, которые в норме должны оставаться недифференцированными. В то же время эктопическая экспрессия Cdx-2 в стенке желудка приводит к формированию ткани, аналогичной кишечнику. Cdx-2 представляет собой важный фактор дифференцировки кишечника, однако до настоящего времени его роль в формировании кишечной трубки на ранних стадиях эмбрионального развития точно не установлена.
– Также рекомендуем “Генетическая регуляция формирования кишечной трубки” Оглавление темы “Эмбриогенез желудочно-кишечного тракта”:
|
Источник