Лактозный оперон кишечной палочки

Лактозный оперон (англ. lac operon) – полицистронный оперон бактерий, кодирующий гены метаболизма лактозы.
Регуляция экспрессии генов метаболизма лактозы у кишечной палочки (Escherichia coli) была впервые описана в 1961 году учеными Ф. Жакобом и Ж. Моно[1] (получившими в 1965 году Нобелевскую премию совместно с А. Львовым). Бактериальная клетка синтезирует ферменты, принимающие участие в метаболизме лактозы, лишь в том случае, когда лактоза присутствует в окружающей среде и клетка испытывает недостаток глюкозы.
Структура
Лактозный оперон (lac operon) состоит из трех структурных генов, промотора, оператора и терминатора. Принимается, что в состав оперона входит также ген-регулятор, который кодирует белок-репрессор.
Структурные гены лактозного оперона – lacZ, lacY и lacA:
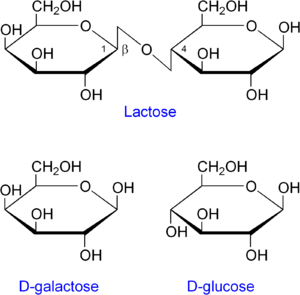
- lacZ кодирует фермент β-галактозидазу, которая расщепляет дисахарид лактозу на глюкозу и галактозу,
- lacY кодирует β-галактозидпермеазу, мембранный транспортный белок, который переносит лактозу внутрь клетки.
- lacA кодирует β-галактозидтрансацетилазу, фермент, переносящий ацетильную группу от ацетил-КoA на бета-галактозиды.
Для катаболизма лактозы необходимы только продукты генов lacZ и lacY, роль продукта гена lacA не ясна. Возможно, что реакция ацетилирования дает бактериям преимущество при росте в присутствии определённых неметаболизируемых аналогов бета-галактозидов, поскольку эта модификация ведет к их детоксикации и выведению из клетки.
Регуляция

Случай, когда есть глюкоза и нет лактозы

Случай, когда есть глюкоза и есть лактоза
Случай, когда нет глюкозы и нет лактозы

Случай, когда нет глюкозы и есть лактоза
РНК-полимераза начинает транскрипцию с промоторного района, который перекрывается с операторным районом. В отсутствие или при низкой концентрации лактозы в клетке белок-репрессор, который является продуктом моноцистронного оперона LacI, обратимо соединяется с операторным районом и препятствует транскрипции. Таким образом, в отсутствие лактозы в клетке ферменты для метаболизма лактозы не синтезируются.
Даже в случае, когда в плазматической мембране клетки отсутствует фермент β-галактозидпермеаза, лактоза из окружающей среды может попадать в клетку в небольших количествах. В клетке две молекулы лактозы связываются с белком-репрессором, что приводит к изменению его конформации и далее к диссоциации белка-репрессора от операторного участка. Может осуществляться транскрипция генов лактозного оперона. При снижении концентрации лактозы новые порции белка-репрессора взаимодействуют с операторными последовательностями и препятствуют транскрипции. Данный механизм регуляции активности лактозного оперона называют негативной индукцией. Веществом-индуктором служит лактоза; при её связывании с белком-репрессором происходит его диссоциация от операторного участка.
Если в клетке концентрация глюкозы достаточная для поддержания метаболизма, активация лактозного оперона не происходит. Промоторная последовательность лактозного оперона «слабая», поэтому даже при отсутствии белка-репрессора на операторном участке транскрипция практически не инициируется. Когда концентрация глюкозы в клетке снижается, происходит активация фермента аденилатциклазы, которая катализирует превращение АТФ в циклическую форму – цАМФ (циклическую форму АМФ в данном случае также называют «сигналом клеточного голода»). Глюкоза является ингибитором фермента аденилатциклазы и активирует фосфодиэстеразу – фермент, катализирующий превращение молекулы цАМФ в АМФ. цАМФ соединяется с белком, активирующим катаболизм (англ. САР, catabolism activating protein), при этом образуется комплекс, который взаимодействует с промотором лактозного оперона, изменяет его конформацию и приводит к повышению сродства РНК-полимеразы к данному участку. В присутствии лактозы происходит экспрессия генов оперона. Белок CAP оказывает положительный контроль на лактозный оперон.
Итак, ферменты для усвоения лактозы синтезируются в клетке кишечной палочки при двух условиях: 1) наличие лактозы; 2) отсутствие глюкозы. Регуляция работы лактозного оперона в зависимости от концентрации лактозы происходит по принципу отрицательной обратной связи: чем больше лактозы – тем больше ферментов для её катаболизма (положительная прямая связь); чем больше ферментов – тем меньше лактозы, чем меньше лактозы – тем меньше производится ферментов (двойная отрицательная обратная связь).
Биологический смысл
Благодаря описанному механизму регуляции транскрипции генов, входящих в состав лактозного оперона, бактерии оптимизируют энергетические затраты, синтезируя ферменты метаболизма лактозы не постоянно, а лишь тогда, когда клетке это необходимо. Сходный механизм регуляции имеется у большинства прокариот; у эукариот он устроен значительно сложнее.
См. также
- Факторы транскрипции
- Лактоза
- Кишечная палочка
Примечания
- ↑ Jacob F; Monod J (June 1961). “Genetic regulatory mechanisms in the synthesis of proteins”. J Mol Biol. 3: 318-56. PMID 13718526.
Ссылки
- Анимация «Регуляция работы лактозного оперона» (англ.)
Источник
Лактозный оперон (lac оперон) – полицистронный оперон бактерий, кодирующий гены метаболизма лактозы.
Регуляция экспрессии генов метаболизма лактозы у кишечной палочки (Escherichia coli) была впервые описана в 1961 году учеными Ф. Жакобом и Ж. Моно[1] (получившими в 1965 году Нобелевскую премию совместно с А. Львовым). Бактериальная клетка синтезирует ферменты, принимающие участие в метаболизме лактозы, лишь в том случае, когда лактоза присутствует в окружающей среде и клетка испытывает недостаток глюкозы.
Структура
Лактозный оперон (lac operon) состоит из трех структурных генов, промотора, оператора и терминатора. Принимается, что в состав оперона входит также ген-регулятор, который кодирует белок-репрессор.
Структурные гены лактозного оперона – lacZ, lacY и lacA:
- lacZ кодирует фермент β-галактозидазу, которая расщепляет дисахарид лактозу на глюкозу и галактозу,
- lacY кодирует β-галактозид пермеазу, мембранный транспортный белок, который переносит лактозу внутрь клетки.
- lacA кодирует β-галактозид трансацетилазу, фермент, переносящий ацетильную группу от ацетил-КoA на бета-галактозиды.
Для катаболизма лактозы необходимы только продукты генов lacZ и lacY, роль продукта гена lacA не ясна. Возможно, что реакция ацетилирования дает бактериям преимущество при росте в присутствии определенных неметаболизируемых аналогов бета-галактозидов, поскольку эта модификация ведет к их детоксикации и выведению из клетки.
Регуляция
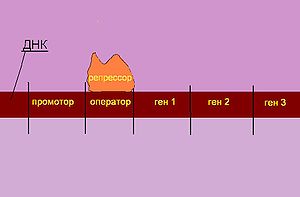
Случай, когда есть глюкоза и нет лактозы

Случай, когда есть глюкоза и есть лактоза

Случай, когда нет глюкозы и нет лактозы

Случай, когда нет глюкозы и есть лактоза
РНК-полимераза начинает транскрипцию с промоторного района, который перекрывается с операторным районом. В отсутствие или при низкой концентрации лактозы в клетке белок-репрессор, который является продуктом моноцистронного оперона LacI, обратимо соединяется с операторным районом и препятствует транскрипции. Таким образом, в отсутствие лактозы в клетке ферменты для метаболизма лактозы не синтезируются.
Даже в случае, когда в плазматической мембране клетки отсутствует фермент β-галактозидпермеаза, лактоза из окружающей среды может попадать в клетку в небольших количествах. В клетке две молекулы лактозы связываются с белком-репрессором, что приводит к изменению его конформации и далее к диссоциации белка-репрессора от операторного участка. Может осуществляться транскрипция генов лактозного оперона. При снижении концентрации лактозы новые порции белка-репрессора взаимодействуют с операторными последовательностями и препятствуют транскрипции. Данный механизм регуляции активности лактозного оперона называют позитивной индукцией. Веществом-индуктором служит лактоза; при её связывании с белком-репрессором происходит его диссоциация от операторного участка.
Если в клетке концентрация глюкозы достаточная для поддержания метаболизма, активация лактозного оперона не происходит. Промоторная последовательность лактозного оперона «слабая», поэтому даже при отсутствии белка-репрессора на операторном участке транскрипция практически не инициируется. Когда концентрация глюкозы в клетке снижается, происходит активация фермента аденилатциклазы, которая катализирует превращение АТФ в циклическую форму – цАМФ (циклическую форму АМФ в данном случае также называют «сигналом клеточного голода»). Глюкоза является ингибитором фермента аденилатциклазы и активирует фосфодиэстеразу – фермент, катализирующий превращение молекулы цАМФ в АМФ. цАМФ соединяется с белком, активирующим катаболизм (англ. САР, catabolism activating protein), при этом образуется комплекс, который взаимодействует с промотором лактозного оперона, изменяет его конформацию и приводит к повышению сродства РНК-полимеразы к данному участку. В присутствии лактозы происходит экспрессия генов оперона. Данный механизм регуляции активности лактозного оперона называют негативной индукцией. «Негативным индуктором» служит глюкоза, которая подавляет активность лактозного оперона.
Итак, ферменты для усвоения лактозы синтезируются в клетке кишечной палочки при двух условиях: 1) наличие лактозы; 2) отсутствие глюкозы. Регуляция работы лактозного оперона в зависимости от концентрации лактозы происходит по принципу отрицательной обратной связи: чем больше лактозы – тем больше ферментов для её катаболизма (положительная прямая связь); чем больше ферментов – тем меньше лактозы, чем меньше лактозы – тем меньше производится ферментов (двойная отрицательная обратная связь).
Биологический смысл
Благодаря описанному механизму регуляции транскрипции генов, входящих в состав лактозного оперона, бактерии оптимизируют энергетические затраты, синтезируя ферменты метаболизма лактозы не постоянно, а лишь тогда, когда клетке это необходимо. Сходный механизм регуляции имеется у большинства прокариот; у эукариот он устроен значительно сложнее.
См. также
- Факторы транскрипции
- Лактоза
- Кишечная палочка
Примечания
- ↑ Jacob F; Monod J (June 1961). «Genetic regulatory mechanisms in the synthesis of proteins». J Mol Biol. 3: 318-56. PMID 13718526.
Ссылки
- Анимация «Регуляция работы лактозного оперона» (англ.)
Источник
Лактозный оперон (англ. lac operon) – полицистронный оперон бактерий, кодирующий гены метаболизма лактозы.
Регуляция экспрессии генов метаболизма лактозы у кишечной палочки (Escherichia coli) была впервые описана в 1961 году учеными Ф. Жакобом и Ж. Моно[1] (получившими в 1965 году Нобелевскую премию совместно с А. Львовым). Бактериальная клетка синтезирует ферменты, принимающие участие в метаболизме лактозы, лишь в том случае, когда лактоза присутствует в окружающей среде и клетка испытывает недостаток глюкозы.
Структура
Лактозный оперон (lac operon) состоит из трех структурных генов, промотора, оператора и терминатора. Иногда принимается, что в состав оперона входит также ген-регулятор, который кодирует белок-репрессор (хотя он находится в другом участке генома и не имеет общего с лактозным опероном промотора).
Структурные гены лактозного оперона – lacZ, lacY и lacA:
- lacZ кодирует фермент β-галактозидазу, которая расщепляет дисахарид лактозу на глюкозу и галактозу,
- lacY кодирует β-галактозидпермеазу, мембранный транспортный белок, который переносит лактозу внутрь клетки.
- lacA кодирует β-галактозидтрансацетилазу, фермент, переносящий ацетильную группу от ацетил-КoA на бета-галактозиды.
Для катаболизма лактозы необходимы только продукты генов lacZ и lacY; роль продукта гена lacA не ясна. Возможно, что реакция ацетилирования дает бактериям преимущество при росте в присутствии определённых неметаболизируемых аналогов бета-галактозидов, поскольку эта модификация ведет к их детоксикации и выведению из клетки.
Регуляция
Случай, когда есть глюкоза и нет лактозы
Случай, когда есть глюкоза и есть лактоза
Случай, когда нет глюкозы и нет лактозы
Случай, когда нет глюкозы и есть лактоза
РНК-полимераза начинает транскрипцию с промоторного района, который перекрывается с операторным районом. В отсутствие или при низкой концентрации лактозы в клетке белок-репрессор, который является продуктом моноцистронного оперона LacI, обратимо соединяется с операторным районом и препятствует транскрипции. Таким образом, в отсутствие лактозы в клетке ферменты для метаболизма лактозы не синтезируются.
Даже в случае, когда в плазматической мембране клетки отсутствует фермент β-галактозидпермеаза, лактоза из окружающей среды может попадать в клетку в небольших количествах. В клетке две молекулы лактозы связываются с белком-репрессором, что приводит к изменению его конформации и далее к диссоциации белка-репрессора от операторного участка. Может осуществляться транскрипция генов лактозного оперона. При снижении концентрации лактозы новые порции белка-репрессора взаимодействуют с операторными последовательностями и препятствуют транскрипции. Данный механизм регуляции активности лактозного оперона называют негативной индукцией. Веществом-индуктором служит лактоза; при её связывании с белком-репрессором происходит его диссоциация от операторного участка.
Если в клетке концентрация глюкозы достаточна для поддержания метаболизма, активация лактозного оперона не происходит. Промоторная последовательность лактозного оперона «слабая», поэтому даже при отсутствии белка-репрессора на операторном участке транскрипция практически не инициируется. Когда концентрация глюкозы в клетке снижается, происходит активация фермента аденилатциклазы, которая катализирует превращение АТФ в циклическую форму – цАМФ (циклическую форму АМФ в данном случае также называют «сигналом клеточного голода»). Глюкоза является ингибитором фермента аденилатциклазы и активирует фосфодиэстеразу – фермент, катализирующий превращение молекулы цАМФ в АМФ. цАМФ соединяется с белком, активирующим катаболизм (англ. САР, catabolism activating protein), при этом образуется комплекс, который взаимодействует с промотором лактозного оперона, изменяет его конформацию и приводит к повышению сродства РНК-полимеразы к данному участку. В присутствии лактозы происходит экспрессия генов оперона. Белок CAP оказывает положительный контроль на лактозный оперон.
Итак, ферменты для усвоения лактозы синтезируются в клетке кишечной палочки при двух условиях: 1) наличие лактозы; 2) отсутствие глюкозы. Регуляция работы лактозного оперона в зависимости от концентрации лактозы происходит по принципу отрицательной обратной связи: чем больше лактозы – тем больше ферментов для её катаболизма (положительная прямая связь); чем больше ферментов – тем меньше лактозы, чем меньше лактозы – тем меньше производится ферментов (двойная отрицательная обратная связь).
Биологический смысл
Благодаря описанному механизму регуляции транскрипции генов, входящих в состав лактозного оперона, бактерии оптимизируют энергетические затраты, синтезируя ферменты метаболизма лактозы не постоянно, а лишь тогда, когда клетке это необходимо. Сходный механизм регуляции имеется у большинства прокариот; у эукариот он устроен значительно сложнее.
См. также
- Факторы транскрипции
- Лактоза
- Кишечная палочка
Примечания
Ссылки
- Анимация «Регуляция работы лактозного оперона» (англ.)
Источник
Оперон, обеспечивающий у E.coliспособность к сбраживанию молочного сахара – лактозы, состоит из промотора, оператора и трех структурных генов. Ген lac Z кодирует фермент- генгалактозидазу, катализирующую гидролиз лактозы до глюкозы и галактозы; ген lac Y-галактозидпермеазу, обеспечивающую транспорт различных сахаров, включая лактозу, мелибиозу и рафинозу, в клетку; ген lac А -тиогалактозидтрансацетилазу, роль которой обычно в утилизации лактозы не ясна. Все три белка обычно присутствуют в клеткахЕ.coliв следовых количествах. Однако при выращивании бактерий на среде, в которой единственным источником углерода и энергии служит лактоза, количество указанных ферментов увеличивается в 1000 раз.
Ген-регулятор лактозного (lac) оперона, обозначаемый lac 1, кодирует белок-репрессор. В активной форме это тетрамер, образованный четырьмя копиями продукта гена lac 1 – полипептидами, состоящими из 360 аминокислот. Клетки с мутациями в гене lac 1 конститутивны по синтезу ферментов, кодируемых генами lac Z, Y и A.
Конститутивный синтез продуктов этих генов возможен не только в случае lac 1 -мутаций в гене репрессоре, но и в случае мутаций в операторе, обозначаемых О . Такие мутации всегда цис-доминантны, поскольку, в отличие от гена-репрессора, оператор может влиять на возможность транскрипции структурных генов только тогда, когда он находится непосредственно рядом с промотором. Если в клетке находится индуктор, он конкурирует с оператором за молекулы репрессора, причем репрессор в первую очередь связывается с индуктором.
. Такие мутации всегда цис-доминантны, поскольку, в отличие от гена-репрессора, оператор может влиять на возможность транскрипции структурных генов только тогда, когда он находится непосредственно рядом с промотором. Если в клетке находится индуктор, он конкурирует с оператором за молекулы репрессора, причем репрессор в первую очередь связывается с индуктором.
В качестве индукторов могут служить различные соединения. Лактоза представляет собой индуктор и одновременно субстрат. В нормальных клетках даже в отсутствие индуктора остаточная активность пермеазы и -галактозидазы обеспечивают возможность проникновения в клетку минимального количества лактозы, которая в результате реакции, катализируемой-галактозидазой, переходит в аллолактозу. Последняя связывается с репрессором, обусловливая его отсоединение от оператора, что, в свою очередь, открывает путь РНК-полимеразе для связывания с промотором и транскрипции генов lac Z, Y и A. К соединениям, являющимся только индукторами, но не субстратами, относятся изопропил-D-тиогалактопиранозид (ИПТГ) и тиометил--D-галактопиранозид (ТМГ), часто используемые для исследования регуляции lac-оперона.
Мутации в промоторе в отличие от мутаций в гене-репрессоре и в операторе не влияют на индуцируемость оперона, однако они регулируют уровень его экспрессии, изменяя эффективность присоединения РНК-полимеразы, и тем самым частоту инициации транскрипции lac-оперона.
Наряду с негативной системой регуляции, lac-оперон контролируется и с помощью позитивно действующих элементов. Их обнаружение связано с исследованием феномена Ж. Моно диаусией, суть которого состоит в том, что утилизация лактозы начнется лишь после того, как будет использована вся имеющаяся в среде глюкоза. Этот феномен, как установили Б. Магазаник с соавторами, – одно из проявленийкатаболитной репрессии или глюкозного эффекта, известного еще с 40-х годов и выражающегося в неспособностиE.coli, катаболизировать различные углеводы (лактозу, арабинозу, галактозу и др.) в присутствии глюкозы, как более эффективного источника энергии.
Расшифровать механизм глюкозного эффекта сумели Р. Перлман и А. Пастан, обнаружившие, что транскрипция lac-оперона контролируется двумя элементами: небольшой молекулой-эффектором, циклическим аденозинмонофосфатом (цАМФ) и белком-активатором САР (от первых букв англ. Catabolite fctivator protein- белок-активатор катаболизма), называемым также белком-рецептором цАМФ. У эукариот цАМФ является медиатором действия гормонов. Оказалось, что добавление цАМФ к растущим в среде с глюкозой клеткам E .coliхотя и замедляет скорость их роста, но снимает катаболическую репрессию, обусловливая тем самым возможность экспрессии лактозного оперона в условиях одновременного присутствия в среде лактозы и глюкозы. Позднее была показана обратная зависимость между содержанием в клетке цАМФ и глюкозы: глюкоза подавляет активность фермента, синтезирующего цАМФ из АТФ. Этот фермент, названныйаденилатциклазой, кодируется геномсуа.
В структуре промотора lac-оперона выявлено два сайта связывания. Один из них взаимодействует с РНК – полимеразой, другой – с комплексом САР-цАМФ. Присоединение комплекса САР-цАМФ к своему сайту на промоторе – условие индукции оперона. Следовательно, этот комплекс позитивно контролирует транскрипцию lac-оперона. Белок САР состоит из двух идентичных субъединиц с общей М около 45000, кодируемых геном САР, илиCRP. Мутации в генесарнарушают участок связывания белка с цАМФ, либо расширяют спектр кофакторов, объединение с которыми обеспечивает индукцию ферментов lac-оперона. У некоторых мутантов в генесартаким кофактором наряду с цАМФ может служить и цГМФ.
около 45000, кодируемых геном САР, илиCRP. Мутации в генесарнарушают участок связывания белка с цАМФ, либо расширяют спектр кофакторов, объединение с которыми обеспечивает индукцию ферментов lac-оперона. У некоторых мутантов в генесартаким кофактором наряду с цАМФ может служить и цГМФ.
В норме, то есть в присутствии глюкозы и в отсутствии цАМФ, белок САР не может объединяться с промотором lac-оперона. В свою очередь, РНК-полимераза не способна эффективно связываться с этим промотором, если к нему не присоединен комплекс САР-цАМФ. Некоторые мутации в промоторе обусловливают независимость экспрессии lac – оперона от глюкозного эффекта, снижая сродство промотора к комплексу САР-цАМФ.
Таким образом, транскрипция lac-оперона на самом деле находится под двойным – негативным и позитивным- контролем. Комплекс САР-цАМФ позволяет РНК-полимеразе присоединиться к матричной ДНК до начала транскрипции. Репрессор – продукт гена lac 1-препятствует инициации синтеза иРНК.
В настоящее время расшифрована полная нуклеотидная последовательность регуляторной области lac-оперона, включающая промотор и оператор. Более того, ДЖ. Шапиро и ДЖ. Беквит с соавторами (1969) сумели выделить чистую ДНК этого оперона, включающую фрагмент гена lac1, полностью промоторную и операторную последовательности, ген lac Z , а также фрагмент гена lac Y. Выяснение структурной организации оператора lac-оперона показало, что существенную роль во взаимодействиях мультимерных белков типа lac-репрессора или РНК-полимеразы с ДНК играют симметричные структуры – палиндромы. Оператор lac-оперона состоит из 26 п.н., из которых 14 представляют собой палиндром: в различных цепях они читаются одинаково, но в противоположных направлениях. Палиндром обнаружен и в участке промотора, связывающемся с комплексом САР-цАМФ.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
07.03.201643.04 Mб26Мондор Г. – Неотложная диагностика. Живот (2 тома)Том 2.djvu
- #
- #
- #
- #
Источник