Обмен липидов в желудочно кишечном тракте

Липиды – обязательная
составная часть пищи. В соответствии с
формулами рационального питания
оптимально следующее соотношение
жировых компонентов: растительные жиры
– 30%, животные – 70%. Распределение жирных
кислот: насыщенные – 30%, моноеновые –
60%, полиеновые –10%.
Переваривание и всасывание
Для переваривания
липидов в желудочно-кишечном тракте
необходимы липолитические ферменты.
Липолиз – это гидролиз липидов.
Липолитические ферменты – гидролазы,
катализирующие гидролиз липидов. Липазы
расщепляют триацилглицериды (жиры),
фосфолипазы – фосфолипиды, холестеразы
– эфиры холестерина.
Липолитические
ферменты проявляют максимальную
активность при рН = 7,8–8,2. Обязательным
условием переваривания липидов в
пищеварительном тракте является
эмульгирование. Эмульгаторы – вещества,
понижающие поверхностное натяжение и
препятствующие склеиванию частиц жира.
В организме природными эмульгаторами
являются соли желчных кислот и сывороточный
альбумин.
Липиды с пищей
поступают в ротовую полость, где нет
липолитических ферментов и переваривания
не происходит. В желудке липаза
присутствует, однако рН желудка (1,0–1,5)
не соответствует оптимуму рН липазы
(7,8–8,2), поэтому происходит лишь
незначительный распад жиров (молока,
яичного желтка). Отделом, в котором
переваривается основная часть липидов,
является тонкий кишечник. Поджелудочная
железа и клетки слизистой оболочки
кишечника секретирует липолитические
ферменты, рН кишечника 7,8–8,2, что близко
к оптимуму.
Жиры гидролизуются
липазой на 90–97%, из них 40% расщепляется
на глицерин и жирные кислоты, 50–57% – на
моноглицериды. Оставшиеся жиры либо
всасываются в тонкой кишке (если размер
жировых капель не превышает 0,5 нм), либо
поступают в толстую кишку и затем
выводятся из организма.
Фосфолипиды
гидролизуются набором фосфолипаз,
которые последовательно гидролизуют
молекулу на составные части. Первый
фермент, фосфолипаза А, отщепляет
ненасыщенную жирную кислоту от второго
углеродного атома глицерина, образуется
лизофосфолипид, который также обладает
свойствами эмульгатора.
Далее последовательно
функционируют фосфолипазы В, С и Д. Они
расщепляют молекулы на глицерин, жирную
кислоту, фосфорную кислоту и азотистое
основание:
Переваривание холестерина
Холестерин
существует в свободной и связанной
формах. Свободная форма – собственно
полициклический спирт холестерин.
Связанная форма – эфиры холестерина,
которые и подвергаются гидролизу в
кишечнике. Эфиры холестерина распадаются
по действием холестераз.
Всасывание
Продукты переваривания
липидов подразделяются на водорастворимые
и жирорастворимые компоненты.
Водорастворимые продукты распада
липидов – глицерин, моноацилглицериды,
Н3РО4,
азотистые основания. Они легко проникают
в клетки слизистой оболочки кишечника.
Жирорастворимые компоненты (жирные
кислоты, холестерин) всасываются с
помощью желчных кислот, с которыми они
образуют водорастворимые комплексы.
К желчным
кислотам
относятся холевая, хенодезоксихолевая
и дезоксихолевая кислоты:
Желчные кислоты
синтезируются из холестерина
Для организма
важны холевая и хенодезоксихолевая
кислоты. Биологически активные формы
– парные желчные кислоты – представляют
собой соединения данных кислот с
гликоколом (глицином) и таурином, они
образуются в желчном пузыре. В кишечнике
парные желчные кислоты соединяются в
мицеллы, наружная часть которых –
гидрофильна, внутренняя – гидрофобна.
Жирные кислоты, холестерин и другие
жирорастворимые вещества проникают в
мицеллу. Мицеллы всасываются в клетки
слизистой оболочки кишечника, затем
распадаются, парные желчные кислоты
всасываются в кровь и поступают в печень,
где снова включаются в состав желчи.
Пищеварительные
липолитические ферменты
Липолизом
называется
гидролиз липидов, липолитическими
ферментами —
ферменты, катализирующие гидролиз
липидов. К пищеварительным липолитическим
ферментам относятся липазы желудка,
липаза, фосфолипаза и холестераза
кишечника.
Липаза
желудка —
слабоактивная гидролаза, гидролизует
жиры молока.
Липаза
тонкого кишечника —
гидролитический фермент, катализирующий
гидролиз жиров. Продуктами гидролиза
являются моно-и диацилглицериды,
глицерин, жирные кислоты. Оптимум рН
7,8—8,2. Активатором являются желчные
кислоты. Обязательным условием
переваривания жиров является их
эмульгирование. К природным эмульгаторам
жиров относятся сывороточный альбумин
и желчные кислоты.
Фосфолипазы
—
гидролазы, катализирующие гидролиз
фосфолипидов в тонком кишечнике.
Активатором фосфолипаз и эмульгаторов
фосфолипидов являются желчные кислоты.
К фосфолипазам относятся фосфолипазы
А, В, С, Д, которые последовательно
гидролизуют молекулу фосфолипидов.
Первый фермент –
фосфолипаза А — отщепляет ненасыщенную
жирную кислоту от второго углеродного
атома глицерина, в результате чего
образуется лизофосфолипид, который
обладает свойствами эмульгатора.
Далее последовательно действуют
фосфолипазы В, С и Д, которые расщепляют
молекулы фосфолипида на глицерин, жирную
и фосфорную кислоты и азотистое основание.
Холестераза
относится
к гидролазам. Субстратом для нее являются
эфиры холестерина (сложносвязанная
форма холестерина). Оптимум рН 7,8—8,2.
Активатор — желчные кислоты. Продукты
гидролиза — свободный холестерин и
кислоты.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник
В полости рта липиды подвергаются лишь механической обработке. В желудке имеется небольшое количество липазы, которая гидролизует жиры. Малая активность липазы желудочного сока связана с кислой реакцией содержимого желудка. Кроме того, липаза может влиять только на эмульгированные жиры, в желудке отсутствуют условия для образования эмульсии жира. Только у детей и у моногастричных животных липаза желудочного сока играет важную роль в переваривании липидов.
Кишечник является основным местом переваривания липидов. В двенадцатиперстной кишке на липиды воздействует желчь печени и сок поджелудочной железы, одновременно происходит нейтрализация кишечного содержимого (химуса). Происходит эмульгирование жиров под действием желчных кислот. В состав желчи входят: холевая кислота, дезоксихолевая (3,12 дигидроксихолановая), хенодезоксихолевая (3,7 дигидроксихолановая) кислоты, натриевые соли парных желчных кислот: гликохолевая, гликодезоксихолевая, таурохолевая, тауродезоксихолевая. Они состоят из двух компонентов: холевой и дезоксихолевой кислот, а также глицина и таурина.
дезоксихолевая кислота хенодезоксихолевая кислота
гликохолевая кислота
таурохолевая кислота
Соли желчных кислот хорошо эмульгируют жиры. При этом увеличивается площадь соприкосновения ферментов с жирами и увеличивается действие фермента. Недостаточность синтеза желчных кислот или задержка поступления нарушает эффективность действия ферментов. Жиры, как правило, всасываются после гидролиза, но часть тонко эмульгированных жиров всасывается через стенку кишечника и переходит в лимфу без гидролиза.
Эстеразы разрывают в жирах эфирную связь между, спиртовой группой и карбоксильной группой карбоновых кислот и неорганических кислот (липаза, фосфатазы).
Под действием липазы жиры гидролизуются на глицерин и высшие жирные кислоты. Активность липазы возрастает под действием желчи, т.е. желчь непосредственно активирует липазу. Кроме того, активность липазы увеличивают ионы Са++ вследствие того, что ионы Са++ образуют нерастворимые соли (мыла) с освободившимися жирными кислотами и предотвращают их подавляющее влияние на активность липазы.
Под действием липазы в начале гидролизуются эфирные связи у α и α1 (боковых) углеродных атомов глицерина, затем у β-углеродного атома:
Под действием липазы до 40% триацилглицеридов расщепляются до глицерина и жирных кислот, 50-55% гидролизуется до 2-моноацилглицеринов и 3-10% не гидролизуется и всасываются в виде триацилглицеринов.
Стериды корма расщепляются ферментом холестеролэстеразой до холестерина и высших жирных кислот. Фосфатиды гидролизуются под влиянием фосфолипаз А, A2, С и D. Каждый фермент действует на определенную сложноэфирную связь липида. Точки приложения фосфолипаз представлены на схеме:
Фосфолипазы поджелудочной железы, тканевые фосфолипазы вырабатываются в виде проферментов и активируются трипсином. Фосфолипаза A2 змеиных ядов катализирует отщепление ненасыщенной жирной кислоты в положении 2 фосфоглицеридов. При этом образуются лизолецитины с гемолитическим действием.
фосфотидилхолин лизолецитин
Поэтому при попадании этого яда в кровь происходит сильный гемолиз.. В кишечнике эта опасность устраняется действием фосфолипазы A1, быстро инактивирующей лизофосфатид в результате отщепления от него остатка насыщенной жирной кислоты с превращением его в неактивный глицерофосфохолин.
Лизолецитины в малых концентрациях стимулируют дифференцировку лимфоидных клеток, активность протеинкиназы С, усиливают клеточную пролиферацию.
Коламинфосфатиды и серинфосфатиды расщепляются фосфолипазой А до лизоколаминфосфатидов, лизосеринфосфатидов, которые далее расщепляются фосфолипазой A2. Фосфолипазы С и D гидролизуют связи холина; коламина и серина с фосфорной кислотой и остатка фосфорной кислоты с глицерином.

Всасывание липидов происходит в тонком отделе кишечника. Жирные кислоты с длиной цепи менее 10 углеродных атомов всасываются в неэтерифицированной форме. Для всасывания необходимо присутствие эмульгирующих веществ – желчных кислот и желчи.
Ресинтез жира, характерного для данного организма, происходит в кишечной стенке. Концентрация липидов в крови в течение 3-5 часов после приема корма высокая. Хиломикроны – мелкие частицы жира, образующиеся после всасывания в кишечной стенке, представляют собой липопротеиды, окруженные фосфолипидами и белковой оболочкой, внутри содержат молекулы жира и желчных кислот. Они поступают в печень, где липиды подвергаются промежуточному обмену, а желчные кислоты проходят в желчный пузырь и далее обратно в кишечник (см. рис.9.3 на стр.192). В результате такого кругооборота теряется малое количество желчных кислот. Считают, что молекула желчной кислоты в сутки совершает 4 кругооборота.
Источник
Эмульгирование и гидролиз липидов
Первые два этапа переваривания липидов, эмульгирование и гидролиз, происходят практически одновременно. Вместе с этим, продукты гидролиза не удаляются, а оставаясь в составе липидных капелек, облегчают дальнейшее эмульгирование и работу ферментов.
Переваривание в ротовой полости
У взрослых в ротовой полости переваривание липидов не идет, хотя длительное пережевывание пищи способствует частичному эмульгированию жиров.
Переваривание в желудке
Собственная липаза желудка у взрослого не играет существенной роли в переваривании липидов из-за ее небольшого количества и того, что ее оптимум рН 4,5-5,5. Также влияет отсутствие эмульгированных жиров в обычной пище (кроме молока).
Тем не менее, у взрослых теплая среда и перистальтика желудка вызывает некоторое эмульгирование жиров. При этом даже низко активная липаза расщепляет незначительные количества жира, что важно для дальнейшего переваривания жиров в кишечнике, т.к. наличие хотя бы минимального количества свободных жирных кислот облегчает эмульгирование жиров в двенадцатиперстной кишке и стимулирует секрецию панкреатической липазы.
Переваривание в кишечнике
Под влиянием перистальтики ЖКТ и составных компонентов желчи пищевой жир эмульгируется. Образующиеся при переваривании лизофосфолипиды также являются хорошим поверхностно-активным веществом, поэтому они способствуют дальнейшему эмульгированию пищевых жиров и образованию мицелл. Размер капель такой жировой эмульсии не превышает 0,5 мкм.
Гидролиз эфиров ХС осуществляет холестерол-эстераза панкреатического сока.
|
Роль колипазы в действии липазы |
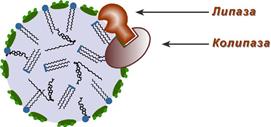
Переваривание ТАГ в кишечнике осуществляется под воздействием панкреатической липазы с оптимумом рН 8,0-9,0. В кишечник она поступает в виде пролипазы, для проявления ее активности требуется колипаза, которая помогает липазе расположиться на поверхности липидной капли.
Колипаза, в свою очередь, активируется трипсином и затем образует с липазой комплекс в соотношении 1:1. Панкреатическая липаза отщепляет жирные кислоты, связанные с С1 и С3 атомами углерода глицерола. В результате ее работы остаются 2-моноацилглицеролы (2-МАГ), которые всасываются или превращаются моноглицерол-изомеразой в 1-МАГ. Последний гидролизуется до глицерола и жирной кислоты. Примерно 3/4 ТАГ после гидролиза остаются в форме 2-МАГ и только 1/4 часть ТАГ гидролизуется полностью.
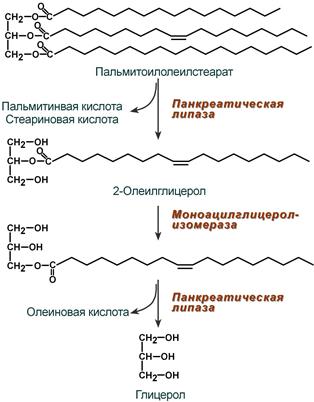
Полный ферментативный гидролиз триацилглицерола
В панкреатическом соке также имеется активируемая трипсином фосфолипаза А2, отщепляющая в фосфолипидах жирную кислоту от С2, также обнаружена активность фосфолипазы С и лизофосфолипазы.
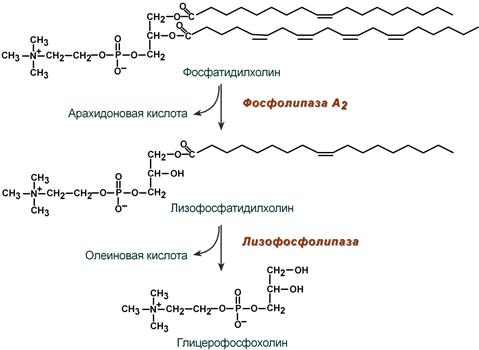
Действие фосфолипазы А2 и лизофосфолипазы на примере фосфатидилхолина
В кишечном соке также имеется активность фосфолипазы А2 и фосфолипазы С.
Для работы всех указанных гидролитических ферментов в кишечнике необходимы ионы Са2+, способствующие удалению жирных кислот из зоны катализа.
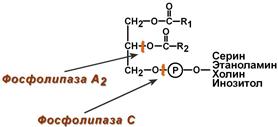
Точки действия фосфолипаз
Образование мицелл
В результате воздействия на эмульгированные жиры ферментов панкреатического и кишечного соков образуются 2-моноацилглицеролы, свободные жирные кислоты и свободный холестерол, формирующие структуры мицеллярного типа (размер уже около 5 нм). Свободный глицерол всасывается напрямую в кровь.
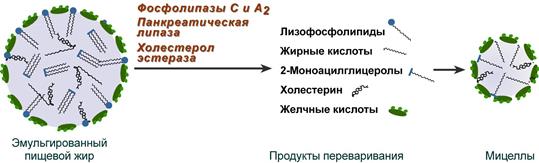
Схематичное изображение переваривания липидов
Полученные смешанные мицеллы достигают эпителия кишечника и их компоненты диффундируют в клетки и попадают в гладкую эндоплазматическую сеть. Желчные кислоты почти не всасываются и остаются в просвете кишечника. Далее желчные кислоты достигают подвздошной кишки и всасываются там (Кишечно-печеночная циркуляция).
Источник
ПЕРЕВАРИВАНИЕ ЛИПИДОВ В ЖЕЛУДОЧНО-КИШЕЧНОМ ТРАКТЕ, СИСТЕМА ИХ ДОСТАВКИ В КЛЕТКИ

Классификация липидов по химическому строению
Простые омыляемые липиды — воска и нейтральные жиры — при щелочном гидролизе дают соли жирных кислот (мыла) и спирты.
Воска — сложные эфиры высших жирных кислот и высших одноатомных спиртов (цетилового, церилового, миристилового).
Высшие жирные кислоты — одноосновные карбоновые кислоты с длинной углеродной цепью, содержащей обычно четное число атомов углерода (от 14 до 24).
Насыщенные — пальмитиновая (16 С), стеариновая (18 С)
Ненасыщенны — олеиновая кислота (18:1 9), линолевая кислота (18:2 6), линоленовая кислота (18:3 3), арахидоновая кислота (20:4 6).
Номенклатура триацилглицеролов, биологическое значение:
1-пальмитоил-2-олеоил-3-стеароилглицерол
Сложные липиды — фосфо-, глико- и сульфолипиды:
Особенности физико-химических свойств — амфифильность.

Гликолипиды — сложные липиды, в состав которых входит аминоспирт сфингозин, аминогруппа которого ацилирована остатком жирной кислоты (церамид), а первичная спиртовая группа связана О-гликозидной связью с углеводами.
В цереброзидах к концевой гидроксильной группе церамида присоединен остаток глюкозы или галактозы. Остаток галактозы некоторых галактоцереброзидов может быть сульфирован по С-3 положению — сульфатиды. В ганглиозидах к церамиду через остаток гексозы присоединен олигосахарид, содержащий хотя бы один остаток нейраминовой кислоты.
Неомыляемые липиды — однокомпонентны и не подвергаются щелочному гидролизу. Стероиды преобладают в липидах животного происхождения, терпены — в липидах растений.
Углеродный скелет терпенов и предшественника стероидов (сквалена) построен из остатков непредельного углеводорода — изопрена:
Терпены представлены терпеновыми углеводородами и их кислородсодержащими производными. Каротиноиды составляют особую группу терпенов; построены из 8 изопреновых единиц (β-каротин).
Высшие спирты:

ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ
Гидролиз жиров пищи в просвете тонкого кишечника. Для последующего всасывания ТАГ сначала должны подвергнуться ферментативному гидролизу до свободных жирных кислот (СЖК) и моноацилглицеролов (МАГ). Гидролиз, хотя и в очень малой степени, начинается в желудке под действием кислой липазы. Всасывание жирных кислот инициирует высвобождение холецистокинина и других биологически активных веществ, которые вызывают выделение в составе панкреатического сока липазы и колипазы, секрецию желчных кислот, которые служат для образования эмульгированного жира и мицелл. Панкреатическая липаза катализирует гидролиз ТАГ. Высвобождающиеся при этом СЖК и МАГ всасываются в двенадцатиперстной кишке и проксимальном отделе тонкого кишечника (около 98 %). Всасывание СЖК осуществляют микроворсинки клеток слизистой. Всасывание холестерола (ХС) происходит менее эффективно, чем ТАГ.
Первичные желчные кислоты синтезируются в печени:

- Конъюгированные формы первичных желчных кислот. Карбоксильная группа боковой цепи желчных кислот может образовывать амидные связи или с глицином, или с таурином. Это обусловливает их эмульгирующие свойства, так как рК ионной группы боковой цепи ниже, чем у исходной карбоксильной группы.

- Вторичные желчные кислоты (дезоксихолевая, литохолевая) образуются в кишечнике под действием ферментов бактерий, которые катализируют отщепление 7-ОН группы и конъюгированной аминокислоты.
Ресинтез липидов в клетках слизистой тонкого кишечника:

ДЕПОНИРОВАНИЕ И МОБИЛИЗАЦИЯ ЛИПИДОВ ИЗ ЖИРОВЫХ ДЕПО,
ВНУТРИКЛЕТОЧНЫЙ ОБМЕН ЖИРНЫХ КИСЛОТ
Состав липопротеинов. Липопротеины состоят из ядра, в котором находятся триацилглицеролы (ТАГ), эфиры холестерола (ЭХ), и поверхностного монослоя из фосфолипидов (ФЛ), свободного или неэстерифицированного холестерола (СХ) и апопротеинов. Функцией липопротеинов является транспорт липидов. Без этой транспортной формы липиды были бы нерастворимы в плазме крови.
Синтез хиломикронов (ХМ). В клетках слизистой кишечника (энтероцитах) идет эстерификация 2-МАГ и ХС жирными кислотами (ЖК), образуются ТАГ и ЭХ, из которых затем формируются ХМ. Всосавшиеся ЖК активируются, преобразуясь в ацил-КоА. Это происходит в гладком ЭР. Важнейшим структурным компонентом ХМ является белок
(апо В-48). В составе одной частицы ХМ находится одна молекула апо В-48.
ХМ секретируются с базолатеральной поверхности клеток кишечника в лимфу, а оттуда, через грудной лимфатический проток, попадают в систему кровообращения. После того как ХМ попадают в лимфу, они получают от ЛПВП апо С-II, С-III и апо Е.
Катаболизм ХМ. Попадая в систему кровообращения, ХМ быстро подвергаются катаболизму. Уровень ТАГ в плазме крови возрастает через 2 ч после приема пищи, а через
4 ч — постепенно снижается. Время разрушения ХМ зависит от гидролиза ТАГ под действием липопротеинлипазы (ЛПЛ). Кофактором этого фермента является апо С-II. Гидролиз ТАГ приводит к уменьшению размеров ХМ, образуется избыточное количество поверхностных элементов по отношению к объему частиц.
Остатки ХМ разрушаются в печени. Таким образом, в процессе своего катаболизма в кровотоке ХМ поставляют ЖК клеткам периферических тканей (жировой и мышечной), в то время как ХС пищи попадает в печень.
Обмен липопротеинов очень низкой плотности (ЛПОНП), липопротеинов промежуточной плотности (ЛППП) и липопротеинов низкой плотности (ЛПНП). Основной функцией этих липопротеинов является транспорт жирных кислот в составе ТАГ из печени к периферическим тканям, особенно в жировую и мышечную. Для синтеза ЛПОНП в гепатоцитах требуется белок апо В-100 и липиды ЭХ, ТАГ и ФЛ.
Триацилглицеролы для ЛПОНП синтезируются путем эстерификации жирных кислот. Они поступают в гепатоциты из плазмы крови (источником их является, например, липолиз в жировой ткани) или синтезируются de novo в печени. Образование ЛПОНП регулируется после трансляции за счет контроля синтеза апо В-100.
Новосинтезированная частица ЛПОНП содержит одну молекулу белка – апо В-100. Другие белковые компоненты, апо С-II, апо С-III и апо Е, поступают на неё от ЛПВП после того, как ЛПОНП попадают в плазму крови. Они требуются для ускорения метаболизма ЛПОНП.
Обмен ЛПОНП. На эндотелии сосудистой стенки ТАГ в составе ЛПОНП подвергаются действию фермента ЛПЛ. Необходимым кофактором для проявления активности ЛПЛ является апо С-II. ЛПЛ образуется в клетках многих тканей, среди которых наибольшее значение имеют жировая ткань, скелетная и сердечная мышцы, молочная железа во время лактации. ЛПЛ катализирует гидролиз ТАГ в составе ХМ и ЛПОНП до жирных кислот, моноацилглицеролов (МАГ), в результате ЛПОНП превращаются в кровотоке в ЛППП. Фермент проявляет слабую активность по отношению к МАГ и ФЛ.
В жировой ткани синтез ЛПЛ стимулирует инсулин. Тем самым обеспечивается поступление жирных кислот в адипоциты для синтеза и хранения в виде ТАГ. В мышцах ЛПЛ позволяет использовать жирные кислоты для окисления в периоды между приемами пищи, а инсулин подавляет образование этого фермента.
Липопротеины промежуточной плотности (ЛППП). Образование ЛППП происходит из ЛПОНП. Около 75 % ЛППП попадает в печень после связывания апо Е с рецепторами для ЛПНП или рецепторами для апо В/апо Е. В печени они полностью разрушаются. Около 25 % ЛППП в кровотоке подвергается действию другого липолитического фермента, печеночной липазы (ПЛ). Этот фермент катализирует дальнейшее расщепление ТАГ в составе ЛППП. В результате ЛППП превращаются в ЛПНП.
Синтез триацилглицеролов. Клетки большинства тканей, особенно печени и жировой ткани, обладают способностью накапливать ТАГ. Жировая ткань функционально специализируется на хранении (запасании) и мобилизации ТАГ.
Предшественниками для синтеза ТАГ являются глицерол-3-фосфат и активированные жирные кислоты. В печени глицерол-3-фосфат может образовываться или в результате фосфорилирования глицерола, или из глюкозы как промежуточный продукт гликолиза. В жировой ткани единственным источником образования глицерол-3-фосфата является гликолиз.
Вслед за перевариванием пищи в плазме крови увеличивается концентрация глюкозы, инсулина, липопротеинов, богатых ТАГ. Стимулируется активность ЛПЛ для гидролиза ТАГ в составе липопротеинов, и снижается активность жиромобилизующей липазы в жировой ткани. Наряду с этим стимулируется образование ТАГ в жировой ткани. Натощак или при повышенной потребности в энергии во время физической работы, повышении уровня катехоламинов, гормона роста, АКТГ и глюкагона в плазме крови, снижении секреции инсулина эти процессы меняются на противоположные – увеличивается липолиз в жировой ткани и высвобождаются жирные кислоты. Они используются в качестве источника энергии. Глицерол используется для глюконеогенеза.
Схема депонирования и расщепления нейтрального жира в адипоцитах:

Липопротеины низкой плотности (ЛПНП). ЛПНП удаляются из кровотока путем взаимодействия с рецепторами для ЛПНП (другое их название апо В/Е рецепторы). Доля этого процесса в удалении всех ЛПНП составляет 75 %. Остальная часть удаляется с помощью «мусорных» рецепторов, имеющих низкую способность связывания. Этот путь получил образное название «мусорный путь».
Поступление ЛПНП в клетки и их внутриклеточный катаболизм:
После связывания ЛПНП комплекс рецептор — ЛПНП переносится в клетку посредством эндоцитоза; затем он сливается с лизосомами и разрушается. Внутриклеточное высвобождение холестерола, происходящее таким путем, вызывает следующие эффекты: а) снижает синтез ключевого фермента образования своего, клеточного, холестерола — ГОМГ-КоА редуктазы; б) снижает синтез рецепторов для ЛПНП; в) активирует АХАТ, который катализирует образование из метаболически активной формы СХ метаболически неактивной формы ЭХ.
Схема реакции, катализируемой АХАТ:

В отличие от регуляторного действия рецепторов к ЛПНП на обмен холестерола в клетках, мусорные рецепторы продолжают транспортировать ХС в клетку без торможения по принципу обратной связи. Тем самым макрофаги превращаются в пенистые клетки. Их образование рассматривается как начальный этап атеросклероза.
Метаболизм липопротеинов высокой плотности (ЛПВП). ЛПВП синтезируются в клетках печени и кишечника. Сразу после секреции ЛПВП имеют вид плоских дисков, содержащих ФЛ. Белковым компонентом их является апо А. Из тканей и клеточных мембран на них поступает холестерол. Под действием фермента лецитин-холестерол ацилтрансферазы (ЛХАТ) из СХ и жирной кислоты фосфатидилхолина образуются ЭХ. В результате частицы ЛПВП созревают, принимая форму глобулы. Затем ЭХ переносятся на ЛППП, ЛПНП, обломки ХМ с помощью липидтранспортирующего белка (ЛПБ) или апо D.
ВНУТРИКЛЕТОЧНЫЙ МЕТАБОЛИЗМ ЖИРНЫХ КИСЛОТ
Жирные кислоты проходят через клеточную мембрану путем диффузии по концентрационному градиенту.
Активация жирных кислот. Первым этапом на пути метаболизма длинноцепочечных жирных кислот в клетке является их активация за счет образования ацил-КоА. Эту реакцию катализирует фермент ацил-КоА синтетаза, который локализован на наружной мембране митохондрий:
Перенос ацил-КоА в митохондрии. Ацил-КоА, имеющий средней длины или короткую углеводородную цепь (<10), может проходить через митохондриальную мембрану путем диффузии. Перенос длинноцепочечного ацил-КоА происходит с помощью карнитин-ацил трансферазы (КАТ). Предшественник для процесса синтеза жирных кислот, малонил-КоА, является аллостерическим ингибитором активности КАТ.
Перенос ацил-КоА в митохондрии:

-ОКИСЛЕНИЕ АЦИЛ-КоА
Последовательность реакций -окисления катализируется 4 ферментами. С их помощью идет дегидрирование, гидратация, образование -кетокислоты и тиолитическое расщепление с высвобождением двухуглеродных фрагментов (ацетил-КоА).
Энергетический выход -окисления на примере пальмитиновой кислоты. Образование АТФ (1,5 АТФ/ФАДН2; 2,5 АТФ/НАДН.H+; 10 АТФ/ацетил-КоА; таким образом, для пальмитоил-КоА (жирная кислота с 16 С): 7 ФАДН2, 7 НАДН.H+ и 8 ацетил-КоА = 108 АТФ).
Расход АТФ на активацию — 1 АТФ (используется энергия гидролиза двух макроэргических связей), в ходе которой пальмитат превращается в пальмитоил-КоА. Чистый энергетический выход для окисления пальмитата — 107 АТФ.
Реакции -окисления жирных кислот:

Окисление ненасыщенных жирных кислот. В ходе -окисления ненасыщенных жирных кислот отщепление двухуглеродных фрагментов ведет к образованию ацил-КоА с двойной связью в цис-положении между С3 и С4. Затем с помощью фермента еноил-КоА изомеразы происходит её превращение в транс-двойную связь между С-2 и С-3. Другой фермент, 2,3-диеноил-КоА редуктаза, может катализировать насыщение двойной связи между С-4 и С-5 в составе ацил-КоА с использованием в качестве кофермента НАДН.Н+. Образовавшийся промежуточный продукт подвергается дальнейшему превращению под влиянием еноил-КоА изомеразы.
ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ В ПЕРОКСИСОМАХ
Окисление жирных кислот в пероксисомах составляет около 30 % всего их окисления. В пероксисомах окисляются необычные жирные кислоты (с длинной углеводородной цепью, дикарбоновые, с разветвленным радикалом). Укорочение радикала в пероксисомах происходит до тех пор, пока не образуется ацил-КоА со средней длиной цепи. Образующийся ацил-КоА с С-8 впоследствии подвергается дальнейшему окислению в митохондриях.
Первоначальная стадия дегидрирования в ходе пероксисомального окисления жирных кислот протекает с образованием Н2О2, а не ФАДН2. Перекись водорода удаляется с помощью каталазы. Все последующие реакции аналогичны происходящим в митохондриях, хотя катализируются они изоферментами пероксисом.
СИНТЕЗ ЖИРНЫХ КИСЛОТ
Субстраты синтеза жирных кислот. Предшественником является ацетил-КоА, процесс протекает в цитозоле. Ацетил-КоА образуется из пирувата под действием митохондриального пируватдегидрогеназного комплекса. Внутренняя митохондриальная мембрана непроницаема для ацетил-КоА. В митохондриях фермент цитратсинтаза катализирует реакцию образования цитрата из ацетил-КоА и ЩУК. Цитрат выходит из митохондрий в цитоплазму. В цитозоле фермент АТФ-цитратлиаза расщепляет цитрат до ацетил-КоА и ЩУК.
Субстраты для синтеза жирных кислот; переход ацетил-КоА из митохондрий в цитозоль:

Ферменты синтеза жирных кислот. В биосинтезе насыщенных жирных кислот участвуют два ферментных комплекса: ацетил-КоА карбоксилаза и ацилсинтетаза.
Ацилсинтетазный мультиферментный комплекс. Комплекс содержит ацилпереносящий белок (АПБ) в качестве своеобразного ядра. Другими ферментами комплекса являются -кетоацилсинтетаза (КС), малонилтрансфераза (МТ), -кетоацил-КоА редуктаза (КР), -гидроксиацилдегидратаза (ГД), еноилредуктаза (ЕР) и ацилтрансацетилаза (АТ).
Ацетил-КоА карбоксилаза катализирует ключевую реакцию в синтезе жирных кислот:
Цитрат активирует фермент. Ацетил-КоА карбоксилаза подвергается обратимому фосфорилированию/дефосфорилированию; цАМФ-зависимая протеинкиназа ингибирует ферментативную активность, а фосфатаза — активирует. На этом основана гормональная регуляция активности ацетил-КоА карбоксилазы. Повышенные концентрации малонил-КоА и ацил-КоА аллостерически ингибируют активность этого фермента.
Схема реакций синтеза жирных кислот:
За 7 таких циклов образуется насыщенная жирная кислота с 16 углеродными атомами. В общем виде это выглядит следующим образом:
ацетил-КоА + 7 малонил-КоА + 14 НАДФН.Н+ + 14 Н+
пальмитат + 7 СО2 + 14 НАДФ + 8 КоА-SH + 6 Н2О
Потребность в НАДФН.Н+ для реакций восстановления в процессе синтеза жирных кислот. Источником восстановленного НАДФ+ является окислительный этап пентозофосфатного пути обмена глюкозы, а также реакции переноса водорода в пути расщепления цитрата. Последние протекают с участием фермента малатдегидрогеназы (кофермент НАДН.Н+) и малик фермента (в реакции восстановления НАДФ+).
ПРОИСХОЖДЕНИЕ НЕНАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТ В КЛЕТКАХ
Незаменимые и заменимые. В организме человека не могут синтезироваться -3 и -6 жирные кислоты в связи с отсутствием ферментной системы, которая могла бы катализировать образование двойной связи в положении -6 или любом другом положении, близко расположенном к -концу. К таким жирным кислотам относятся: линолевая кислота (18:2, 9, 12), линоленовая кислота (18:3, 9, 12, 15), арахидоновая кислота (20:4, 5, 8, 11, 14). Арахидоновая кислота является незаменимой только при недостатке линолевой кислоты.
Функции полиненасыщенных жирных кислот: придают жидкостность мембранам, являются предшественниками эйкозаноидов
Эйкозаноиды (липидные гормоны). Простагландины, тромбоксаны и лейкотриены. Самым главным предшественником является арахидоновая кислота.
Схема высвобождения арахидоновой кислоты из мембранных фосфолипидов:

Судьба арахидоновой кислоты, которая высвободилась из состава мембранных фосфолипидов:

Ингибиторы синтеза эйкозаноидов. Кортикостероиды ингибируют фосфолипазу А2, тем самым они ингибируют высвобождение арахидоновой кислоты из состава мембранных фосфолипидов. Ингибиторы из числа противовоспалительных препаратов нестероидной природы (аспирин, индометацин, фенилбутазон) ингибируют циклоксигеназу.
Простагландины стимулируют сокращение гладких мышц, липолиз, снижают секрецию желудочного сока, свертывание крови, вызывают расширение кровеносных сосудов.
Тромбоксаны стимулируют агрегацию тромбоцитов, сокращение стенки артерий, образование цАМФ в тромбоцитах, повышают кровяное давление.
Лейкотриены участвуют в формировании воспаления, аллергических реакций, в хе