Регуляция функций желудочно кишечного тракта

Оглавление темы “Функция всасывания кишечника. Пищеварение в ротовой полости и функция глотания.”: Гуморальная регуляция секреции пищеварительных соков и моторики желудка и кишечника. Гормональная регуляция пищеварительного тракта.Центральные, периферические и местные рефлексы осуществляются в тесном взаимодействии с гуморальным механизмом регуляции миоцитов, гландулоцитов и нервных клеток. В слизистой оболочке желудочно-кишечного тракта и в поджелудочной железе имеются эндокринные клетки, которые вырабатывают гастроинте-стинальные гормоны (регуляторные пептиды, энтерины). Эти гормоны через кровоток и местно (паракринно, диффундируя через межклеточную жидкость) оказывают влияние на миоциты, гландулоциты, интрамураль-ные нейроны и эндокринные клетки. Их выработка запускается рефлекторно (через блуждающий нерв) во время приема пищи и длительное время поддерживается за счет раздражающего влияния продуктов гидролиза пищевых веществ и экстрактивных веществ. Таблица 11.1. Гормоны желудочно-кишечного тракта, место их образования и вызываемые ими эффекты
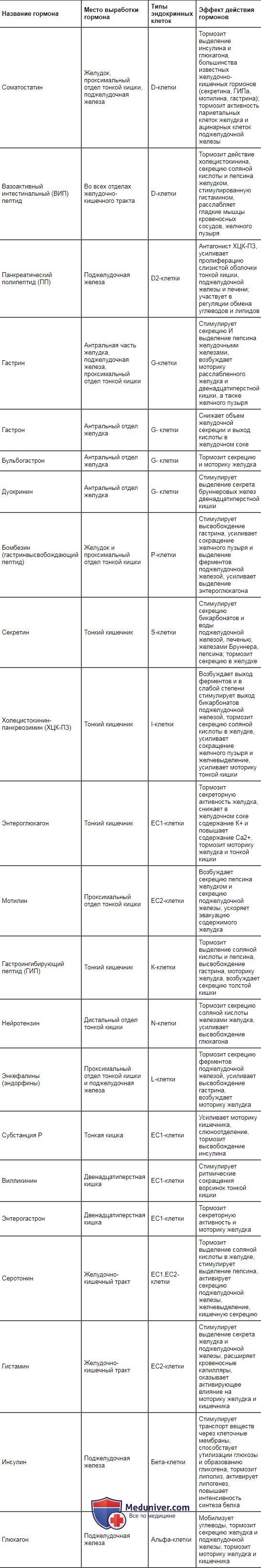 Место выработки основных гастроинтестинальных гормонов, вызываемые ими эффекты и продуцирующие их клетки представлены в табл. 11.1. В настоящее время обнаружено около 30 регуляторных пептидов. Как следует из представленной таблицы, они оказывают стимулирующее, тормозное и модулирующее влияние на секрецию пищеварительных соков, моторику гладкой мускулатуры желудочно-кишечного тракта, всасывание, выделение энтеринов эндокринными элементами слизистой оболочки желудка, кишечника и поджелудочной железы. Выделение гастроинтестинальных гормонов имеет каскадный характер. Например, под влиянием гастрина обкладочные клетки желез желудка увеличивают выработку соляной кислоты, которая в слизистой оболочке тонкой кишки стимулирует выделение S- и J-клетками секретина и холеци-стокинна — панкреозимина. Секретин усиливает секрецию воды и бикарбонатов поджелудочной железой и печенью, а холецистокинин — панкреозимин — возбуждает выделение ферментов поджелудочной железой и тормозит секрецию соляной кислоты обкладочными клетками, усиливает моторику тонкой кишки и желчного пузыря. Регуляторные пептиды, поступая в кровоток, быстро разрушаются в печени и почках и тем самым создают условия для осуществления эффектов других гастроинтестинальных гормонов. Выработка некоторых энтеринов носит циклический характер и может осуществляться и при отсутствии пищевого раздражителя. Например, мо-тилин, вырабатываемый ЕС2-клетками проксимального отдела тонкой кишки, вызывает сокращения мышц желудка и кишечника, совпадающие с периодами «голодной» активности пищеварительного тракта. Видео физиология гуморальной регуляции и ее отличие от гормональной – профессор, д.м.н. П.Е. Умрюхин– Также рекомендуем “Схема механизмов регуляции функций желудочно-кишечного тракта (жкт). Обобщенная схема механизмов регуляции функций пищеварительного тракта.” |
Источник
Регуляция функции желез пищеварительного тракта. Нервная регуляция секреции желез
а) Основные механизмы стимуляции желез пищеварительного тракта. Эффект взаимодействия пищи с эпителием. Функция раздражения энтеральной нервной системы. Механическое раздражение пищей в определенном отделе желудочно-кишечного тракта обычно вызывает адекватную секрецию желез этой области и часто — смежных с ней областей. Часть этих местных эффектов, особенно секреция слизи слизистыми клетками, происходит за счет непосредственного раздражения пищей поверхности железистых клеток.
Кроме того, местная стимуляция эпителия активирует энтеральную нервную систему кишечной стенки. Раздражителями могут служить следующие стимулы:
(1) тактильная стимуляция;
(2) химическое раздражение;
(3) растяжение кишечной стенки.
В результате стимуляции как слизистые клетки эпителиальной поверхности кишки, так и глубокие железы кишечной стенки рефлекторно усиливают свою секрецию.
Фазы желудочной секреции и их регуляция
б) Стимуляция секреции автономной нервной системой. Парасимпатическая стимуляция. Раздражение окончаний парасимпатических нервов пищеварительного тракта, как правило, увеличивает уровень секреции пищеварительных желез. В первую очередь это касается желез верхней части тракта (иннервируемых языкоглоточным и блуждающим нервами), таких как слюнные железы, железы пищевода, железы желудка, а также поджелудочная железа и бруннеровы железы двенадцатиперстной кишки. Это также справедливо для некоторых желез дистального отдела толстой кишки, иннервируемых тазовыми парасимпатическими нервами. Секрецию в остальных отделах тонкой кишки и в первых двух третях толстой кишки осуществляют главным образом местные нервные и гормональные стимулы в каждом отделе пищеварительной трубки.
в) Симпатическая стимуляция. Раздражение симпатических нервов, идущих к желудочно-кишечному тракту, приводит к умеренному локальному увеличению секреции некоторых желез. Но симпатическая стимуляция также вызывает сужение сосудов, которые кровоснабжают железы. В результате симпатическая стимуляция может производить двойной эффект:
(1) отдельная симпатическая стимуляция обычно незначительно увеличивает секрецию;
(2) если парасимпатическая или гормональная стимуляция уже вызвала обильную секрецию, то наслаивающаяся симпатическая стимуляция обычно уменьшает секрецию, иногда значительно, главным образом из-за уменьшения кровоснабжения вследствие сужения сосудов.
г) Гормональная регуляция секреции желез. В желудке и кишечнике несколько различных гастроинтестинальных гормонов помогают регулировать объем и характер секреции. Эти гормоны высвобождаются из слизистой желудочно-кишечного тракта в ответ на присутствие пищи в полости пищеварительного тракта. Затем гормоны всасываются в кровь и достигают желез, где вызывают секрецию. Этот тип стимуляции особенно важен для увеличения выброса желудочного и панкреатического соков, когда пища поступает в желудок и двенадцатиперстную кишку. По своему химическому составу гастроинтестинальные гормоны представляют собой полипептиды либо их производные.
– Также рекомендуем “Секреция воды и электролитов в ЖКТ. Физиология слизи желудочно-кишечного тракта”
Оглавление темы “Желудочная и поджелудочная секреция”:
1. Регуляция функции желез пищеварительного тракта. Нервная регуляция секреции желез
2. Секреция воды и электролитов в ЖКТ. Физиология слизи желудочно-кишечного тракта
3. Физиология слюны. Секреция слюны
4. Гигиенические свойства слюны. Нервная регуляция слюноотделения
5. Секреция в пищеводе. Физиология желудочной секреции
6. Физиология секреции пепсиногена. Секреция желудочной слизи и гастрина
7. Стимуляция секреции кислоты в желудке. Стимуляция секреции гастрина
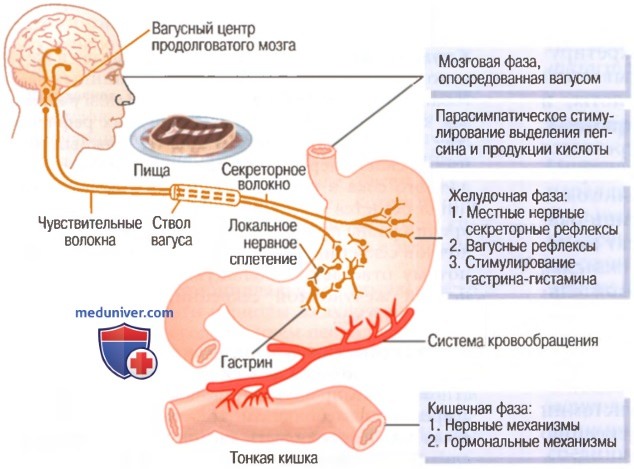
8. Физиология и фазы желудочной секреции. Торможение и регуляция желудочной секреции
9. Строение гастрина. Физиология секреции поджелудочной железы
10. Предотвращение самопереваривания поджелудочной железы. Секреция ионов бикарбоната
Источник
Оглавление темы “Функция всасывания кишечника. Пищеварение в ротовой полости и функция глотания.”: Схема механизмов регуляции функций желудочно-кишечного тракта (жкт). Обобщенная схема механизмов регуляции функций пищеварительного тракта.Обобщенная схема механизмов регуляции функций пищеварительного тракта представлена на рис. 11.9. Эта схема демонстрирует взаимодействие нервных и гуморальных механизмов регуляции миоцитов, гландулоцитов и эндокринных клеток пищеварительного тракта. В ответ на внешние раздражители, адресованные экстерорецепторам, структуры пищеварительного тракта через вегетативные нервные волокна получают регулирующие влияния, причем по парасимпатическим волокнам — возбуждающие, а по симпатическим — тормозные.
При раздражении интерорецепторов слизистой оболочки желудочно-кишечного тракта компонентами химуса (продуктами гидролиза пищи, экстрактивными веществами, крупными фрагментами пищи) рефлекторно изменяется активность мышечных, секреторных и эндокринных элементов. Рефлекс с интерорецепторов удлиняет во времени секреторную и моторную реакцию, вызванную раздражением экс-терорецепторов. Эндокринное звено этого рефлекса подкрепляется ответом эндокриноцитов на непосредственные химические раздражители, диффундирующие в слизистую оболочку пищеварительного тракта из химуса, заполняющего его полость. Гастроинтестинальные гормоны через кровоток и интерстициальные пространства достигают регулируемых секреторных, мышечных и эндокринных элементов. Нервные импульсы от рецепторов желудочно-кишечного тракта по афферентным путям достигают пищевого центра, доставляя информацию о состоянии процесса пищеварения. Она дополняется гуморальными влияниями, обусловленными поступлением в кровь продуктов гидролиза пищевых веществ и гастроинтестинальных гормонов. Информация, поступающая в пищевой центр по каналам нервной и гуморальной обратной связи, вызывает его ответную реакцию, направленную на оптимизацию процесса пищеварения и пищевого поведения животного и человека. – Также рекомендуем “Периодическая деятельность пищеварительной системы. Голодная периодическая деятельность пищеварительного тракта. Мигрирующий моторный комплекс.” |
Источник
Транзит содержимого желудочно-кишечного тракта (ЖКТ) обеспечивается его мышечными элементами и находится под многоуровневой системой контроля, которая может нарушаться при тех или иных патологических состояниях, в том числе при травме спинного мозга.
Двигательная активность ЖКТ включает в себя два вида сокращений: непропульсивные (перемешивающие) движения и пропульсивная перистальтика (превалирование тех или иных сокращений зависит от отдела ЖКТ).
Регуляция моторики ЖКТ осуществляется:
- водителями ритма в структуре гладкомышечных клеток ЖКТ;
- собственной энтеральной (внутренней) нервной системой (ауэрбахово и мейсснерово сплетения);
- вегетативной нервной системой (ВНС);
- гормонами, регуляторными пептидами и биологически активными веществами;
- содержимым просвета пищеварительной трубки (количество, твердость, состав, калорийность, кислотность и др.).
Особенностью гладкомышечных клеток ЖКТ является то, что помимо собственно сократительной активности они обладают способностью спонтанно генерировать электрические импульсы и выступать в роли водителей ритма для других мышечных элементов желудочной и кишечной стенок. Данное свойство лежит в основе регуляции моторной и эвакуаторной функций пищеварительного тракта. В свою очередь, внешние нервные и гормональные воздействия индуцируют и модулируют сокращения и определяют его силу и продолжительность.
Вопрос о локализации «пейсмейкера» («водителя ритма») ЖКТ остается открытым. Имеющиеся данные позволяют предположить, что «водитель ритма» желудка расположен в проксимальной части большой кривизны, а для тонкой кишки данную роль играет проксимальный отдел двенадцатиперстной кишки (некоторые авторы локализуют его в области впадения общего желчного протока). Они генерируют медленные электрические волны наиболее высокой для всей тонкой кишки частоты. Однако также доказано, что любая зона желудочно-кишечного тракта является источником ритма для каудально расположенных сегментов или становится таковым в определенных условиях. Следует отметить, что всегда существует градиент как основного электрического ритма, так и ритмических сокращений гладких мышц желудочно-кишечного тракта по частоте и скорости проведения возбуждения в каудальном направлении (орально-анальный градиент). Благодаря наличию этого градиента содержимое ЖКТ медленно продвигается в каудальном направлении даже во время непропульсивной перистальтики.
Все отделы ЖКТ, кроме верхней трети пищевода и наружного кольца анального сфинктера, где имеются поперечнополосатые мышцы, имеют гладкомышечное строение (то есть автономную иннервацию), и человек таким образом лишен возможности сознательной (произвольной) регуляции их состояния.
В регуляции моторной и секреторной деятельности ЖКТ участвуют симпатические и парасимпатические нервные влияния (табл. 1).
Таблица 1. Влияние парасимпатического и симпатического отделов ВНС на моторику ЖКТ
Отдел ЖКТ | Парасимпатический отдел ВНС | Симпатический отдел ВНС |
Пищевод | Уменьшает тонус нижнего пищеводного сфинктера | Замедление перистальтики. Повышение тонуса нижнего пищеводного сфинктера |
Желудок | Усиление перистальтики. Расслабление сфинктера привратника | Замедление перистальтики. Сокращение сфинктера привратника |
Желчный пузырь | Усиление перистальтики | Замедление перистальтики |
Тонкая кишка | Усиление перистальтики | Замедление перистальтики |
Толстая кишка | Усиление перистальтики. Расслабление внутреннего сфинктера прямой кишки | Замедление перистальтики. Сокращение внутреннего сфинктера прямой кишки |
Ядра парасимпатических нервов расположены в среднем и продолговатом мозге и в сером веществе боковых рогов крестцового отдела спинного мозга (SII-IV). Ядра краниальной локализации дают начало, в частности, блуждающему нерву (Х-пара; n. vagus), обеспечивающему парасимпатической иннервацией практически весь ЖКТ, за исключением дистальных его отделов.
Парасимпатические нервные волокна оканчиваются в ганглиях интрамуральных сплетений или в ганглиях, расположенных в стенках слюнных желез и печени. Нейромедиатором в преганглионарных и постганглионарных нервах служит ацетилхолин, хотя существует множество биологически активных пептидов, также играющих роль постганглионарных медиаторов: вазоактивный интестинальный полипептид (VIР),энкефалины, вещество Р(SР), серотонин и др.
Парасимпатические волокна стимулируют перистальтику желудка и ускоряют время эвакуации из него пищи.
Симпатические нервы берут начало в боковых рогах спинного мозга, тораколюмбального его отдела (CVIII, ThI – LIII), и оканчиваются в чревном ганглии (для пищевода, желудка, двенадцатиперстной кишки, поджелудочной железы), верхнем брыжеечном ганглии (для тонкой кишки и верхней части толстой кишки) и нижнем брыжеечном ганглии (для нижнего отдела толстой кишки и анального отверстия). Нейромедиатором в преганглионарных волокнах служит ацетилхолин, а в постганглионарных – норадреналин.
Каждый отдел ВНС содержит также висцеральные афферентные волокна, сигналы по которым поступают в центральную нервную систему (ЦНС), участвуя в запуске безусловных рефлексов и возникновении ощущений.
Существует разница в активации продольных и циркулярных мышц. Продольные мышцы иннервируются холинергическими волокнами и находятся в сокращенном состоянии до тех пор, пока действует стимул. В циркулярных мышцах в ответ на нервный импульс сначала развивается небольшое кратковременное напряжение и только после окончания действия стимула начинается сокращение. Предполагают, что нейромедиатором в циркулярных мышцах служит скорее всего ВИП (вазоактивный интестинальный пептид).
К энтеральной нервной системе относится нервная сеть, состоящая из двух отделов:
— межмышечное (ауэрбахово) сплетение, лежащее между слоями продольных и циркулярных мышц;
— подслизистое (мейсснерово) сплетение, находящееся между слоем циркулярных мышц и подслизистым мышечным слоем.
Ауэрбахово сплетение регулирует тонус гладких мышц ЖКТ и ритм их сокращений, участвуя также в регуляции процессов секреции и всасывания. Мейсснерово сплетение регулирует секреторную активность эпителиальных клеток. Афферентные волокна обоих сплетений передают сенсорные сигналы от расположенных в органах ЖКТ рецепторов в ЦНС.
В состав внутренних сплетений ЖКТ входят три функциональных типа клеток:
— внутренние афферентные нейроны, содержащие ацетилхолин или другие нейромедиаторы, в частности SP;
— интернейроны, образующие нейронную сеть и обеспечивающие взаимодействие между различными участками сплетения;
— мотонейроны, которые, в свою очередь, подразделяются на стимулирующие (содержащие ацетилхолин и SP) и ингибирующие (содержащие в качестве медиаторов NO и VIР).
Значение данного «отдела» нервной системы в регуляции моторики ЖКТ хорошо видно на примере болезни Гиршпрунга, когда отсутствие межмышечного сплетения в определенном участке толстой кишки приводит к состоянию постоянного тонического сокращения данного участка.
Благодаря внутренней нервной системе ЖКТ может функционировать и в полном «отрыве» от ЦНС, однако его работа и адаптация будет эффективнее при сохраненных связях со спинным мозгом, а координация произвольной и автономной активности, естественно, невозможна без связи с головным мозгом.
В настоящее время в слизистой ЖКТ и в поджелудочной железе обнаружено по меньшей мере 18 видов клеток, вырабатывающих различные гормоны или пептиды, участвующие в регуляции функций ЖКТ (табл. 2).
Таблица 2. Основные регуляторные пептиды и их влияние на моторную функцию различных отделов ЖКТ
Регуляторные пептиды | Основные функции |
Гастрин | Повышает давление в области верхнего пищеводного сфинктера. Замедляет эвакуацию пищи из желудка. Усиливает моторику привратника. Усиливает моторику толстой кишки |
Секретин | Снижает давление в области верхнего пищеводного сфинктера. Подавляет перистальтику желудка. Угнетает моторику толстой кишки |
Холецистокинин | Снижает давление в области верхнего пищеводного сфинктера. Усиливает перистальтику желудка. Увеличивает эластичность желудка. Стимулирует сокращения желчного пузыря. Усиливает моторику толстой кишки |
Соматостатин | Подавляет перистальтику желудка |
Мотилин | Повышает давление в области вехнего пищеводного сфинктера. Усиливает моторику желудка |
Вилликинин | Стимулирует сокращение ворсинок тонкой кишки |
Желудочно-ингибирующий пептид | Угнетает моторику желудка |
Панкреатический полипептид | Повышает давление в области верхнего пищеводного сфинктера |
Энтероглюкагон | Снижает давление в области верхнего пищеводного сфинктера. Подавляет перистальтику желудка. Стимулирует желчеотделение. Угнетает моторику толстой кишки |
Нейротензин | Угнетает секрецию и опорожнение желудка, вызывает сужение сосудов |
Глюкозозависимый инсулинотропный пептид | Вызывает высвобождение инсулина. Снижает давление в области верхнего пищеводного сфинктера |
Вазоактивный интестинальный полипептид | Снижает давление в области вехнего пищеводного сфинктера. Подавляет перистальтику желудка. Расслабляет гладкие мышцы |
Вещество Р | Стимулирует сокращение гладких мышц. Повышает давление в области верхнего пищеводного сфинктера |
Энкефалины, эндорфины | Угнетают сокращения гладких мышц |
Регуляция образования гормоноподобных веществ в ЖКТ отличается от таковой в других эндокринных системах тем, что их секреция зависит не столько от концентрации пептидов в крови, сколько от прямого взаимодействия компонентов желудочного или кишечного содержимого с эндокринными клетками пищеварительного тракта.
Характеристики химуса (количество, твердость, состав, кислотность, калорийность, размер частиц и т.д.) играют важную роль в регуляции моторики ЖКТ. Так, например, во время фазы пищеварительного опорожнения из желудка могут выводиться только частицы размером не более 3-5 мм, частицы, размеры которых превышают указанные цифры, попадают в двенадцатиперстную кишку только в голодную фазу, благодаря существованию «голодной» перистальтической активности. Давление в области нижнего пищеводного сфинктера повышается после приема белковой пищи, кислое содержимое эвакуируется из желудка медленнее, чем нейтральное, гиперосмолярное содержимое – медленнее, чем гипоосмолярное, а липиды – медленнее, чем продукты расщепления белков. Похожая зависимость существует и при пассаже химуса по тонкой кишке: наибольшую скорость перемещения имеют углеводы, а наименьшую жиры. Обратные влияния наблюдаются в толстой кишке, где жиры усиливают моторику, а углеводы и белки на нее не влияют, по крайней мере напрямую.
Особые механизмы лежат в процессах перемещения пищи в начальном и конечном участках пищеварительной системы: в актах глотания и дефекации. Особенностью является участие поперечнополосатой мускулатуры и возможность осознанной регуляции указанных процессов. В то же время это различие в итоге оказывается достаточно условным, так как психическая активность может оказывать влияние не только на осознанные, но также и на неосознанные процессы, такие, как моторика кишечника, опосредованно через ВНС.
Дефекация является сложным скоординированным рефлекторным актом, в котором принимают участие кора головного мозга, ВНС, рецепторы прямой кишки, мускулатура брюшного пресса и толстой кишки. Если на первом году жизни ребенка дефекация протекает по типу безусловного рефлекса, то со второго года жизни начинают устанавливаться условно-рефлекторные связи на время суток (утро, вечер), место (туалетная комната, детский горшок), звук и многое др.
Важным феноменом, обеспечивающим контроль над эвакуацией кишечного содержимого, является так называемое держание. Оно осуществляется функциональной системой, состоящей из промежностного отдела прямой кишки с внутренним сфинктером, наружного сфинктера и мышцы, поднимающей задний проход. Различают два типа держания: кишечное и анальное, представляющие собой последовательные этапы единого механизма опорожнения кишечника.
Кишечное держание обеспечивает продвижение содержимого по кишке в течение более-менее длительного периода времени, чему способствуют форма толстой кишки в виде обода, гаустрация, координированные антиперистальтические движения, ректосигмоидный изгиб, сдерживающий продвижение кишечного содержимого к прямой кишке. Данный этап держания является неосознанным. Анальное держание осуществляется за счет рефлекторного и произвольного сокращения запирательного аппарата прямой кишки: внутреннего и наружного сфинктеров. При этом внутренний сфинктер играет роль постоянного пассивного замыкателя, обеспечивая тоническое смыкание стенок заднепроходного канала, а активное сокращение замыкательного аппарата достигается при помощи наружного сфинктера.
Действие наружного сфинктера (по A. Shank, 1975) реализуется с помощью трех петель: верхней, средней и нижней. Верхнюю петлю образует так называемое аноректальное кольцо, фиксирующееся к внутренней нижней поверхности лонной кости, среднюю — поверхностная порция наружного сфинктера, прикрепляющаяся к копчику, а нижнюю — подкожная порция этой мышцы, прикрепленная к перианальной коже в области срединного шва промежности. Верхняя и нижняя петли, иннервируемые геморроидальными ветвями полового нерва, перетягивают заднюю анальную стенку кпереди, в то время как средняя петля, иннервируемая четвертым крестцовым нервом, тянет переднюю анальную стенку кзади. Названные петли являются, по существу, отдельными сфинктерами и, взаимодействуя, обеспечивают полное держание кишечного содержимого.
Перистальтику и функцию запирательного аппарата прямой кишки регулируют главным образом три нервных центра:
1) ганглии стенки прямой кишки и нижнее подчревное сплетение (plexus hypogastricus inf.);
2) спинномозговой центр на уровне SII—IV;
3) кора головного мозга.
Регуляция функции осуществляется тремя рефлекторными дугами. Первая дуга регулирует расслабление внутреннего сфинктера и перистальтические движения ректоанального сегмента через интрамуральный путь и тазовые сплетения, в частности подчревные. Нейроны спинномозгового центра координируют перистальтические движения и контролируют непроизвольное действие сфинктерного аппарата.
Вторую дугу образуют афферентный путь, проводящий импульсы от ректальных рецепторов к нейронам спинномозгового центра, и эфферентный путь, проводящий импульсы от спинномозгового центра к лонно-прямокишечной мышце и наружному сфинктеру. Этот рефлекс возникает при повышении давления в прямой кишке или при растяжении ее стенки и вызывает сокращение лонно-прямокишечной мышцы и наружного сфинктера, таким образом обеспечивая держание кала.
Через третью дугу кора головного мозга управляет функцией наружного сфинктера и мышц, поднимающих задний проход. Афферентные импульсы от барорецепторов леваторной мышцы через половой нерв и спинной мозг достигают коры головного мозга, и оттуда эфферентные импульсы через крестцовый спинномозговой центр возвращаются к наружному сфинктеру и леваторной мышце. По этой дуге чувство позыва к дефекации передается к коре головного мозга, и при помощи произвольного сокращения или расслабления наружного сфинктера и леваторной мышцы обеспечиваются условия для держания или дефекации.
Если держание кишечного содержимого — относительно пассивный процесс и может осуществляться непроизвольно за счет самоуправления ректального и спинномозгового центров, то физиологическая дефекация является активным процессом и осуществляется под контролем сознания. Акт дефекации состоит из двух фаз. В первой, непроизвольной, кишечное содержимое поступает пропульсивной волной в прямую кишку, при растяжении которой расслабляется внутренний сфинктер, давление на его уровне становится ниже, чем в прямой кишке, и кишечное содержимое продвигается дистальнее до входа в анальный канал, вступая в контакте чувствительной зоной его слизистой оболочки. Вскоре тонус сфинктера восстанавливается, а кишечное содержимое остается выше анального канала.
В первой фазе держание обеспечивается за счет рефлекторного сокращения (по второй дуге) наружного сфинктера и лонно-прямокишечной мышцы. Последняя, перетягивая кишку кзади, закрывает вход в анальный канал. В это время перистальтика усиливается, кишечное содержимое поступает в ампулу, последняя еще больше растягивается; раздражаются не только рецепторы ампулы прямой кишки, но и леваторные мышцы. Импульсы достигают коры головного мозга по третьей рефлекторной дуге, возникает позыв к дефекации и наступает следующая, произвольная, фаза. Если условия для этого социально приемлемы, то при анализе чувства позыва кора головного мозга подает команду совершить акт дефекации. Наружный сфинктер произвольно расслабляется, увеличивается аноректальный угол, внутренний сфинктер по?