В процессе дифференцировки из кишечной энтодермы образуется

Регуляция специализации энтодермы при формировании желудочно-кишечного тракта у плодаРазвитие ЖКТ происходит в три этапа. Первый этап — первичная специализация энтодермы. Второй этап заключается в формировании и структурировании кишечной трубки, что определяет передне-заднюю ось эмбриона и границы между различными органами. На третьем этапе начинается образование органов, которые в процессе развития выходят за границы кишечной трубки, например печени и поджелудочной железы. В экспериментах на лабораторных животных установлены эволюционно консервативные группы генов, принимающих участие в специализации энтодермы, и ряд других генов, участвующих только в процессе развития ЖКТ позвоночных. Эпителий этого тракта формируется из энтодермы — одного из эмбриональных зародышевых листков, возникающих в процессе гаструляции. Исследования, проведенные на таких лабораторных организмах, как круглые черви (почвенные нематоды Caenorhabdilis elegans), плодовые мушки (род Drosophila), африканские шпорцевые лягушки (род Xenopus), аквариумные рыбки Danio rerio и лабораторные мыши, позволили установить некоторые основные молекулярные механизмы регуляции процесса развития энтодермы. Моделирование на лабораторных мышах, у которых был проведен нокаут определенных генов для исследования их функций, в настоящее время рассматривается в качестве основы для изучения таких заболеваний ЖКТ у человека, как стеноз привратника, различные атрезии, неперфорированный анус. Специализация энтодермы может быть прослежена на самых ранних стадиях развития эмбриона. В классических экспериментальных исследованиях продемонстрирована способность эксплантатов куриных эмбрионов еще до процесса гаструляции, в культуре, к развитию ЖКТ, что свидетельствует о четкой детерминации процесса уже на данном этапе эмбрионального развития. Энтодерма куриных эмбрионов, выделенная и помещенная в лабораторных условиях в культуру до формирования ЖКТ, характеризуется экспрессией молекулярных маркеров, которые должны в норме появиться in vivo, что свидетельствует о наличии феномена ранней специализации. Появляются доказательства в поддержку гипотезы, что отличительные особенности энтодермы обусловлены функциональной клеточной независимостью, однако для нормального развития органов необходимо тесное взаимодействие энтодермы и мезодермы. В настоящее время на многочисленных лабораторных организмах установлены целые группы генов, способствующих специализации энтодермы. Один класс генов кодирует факторы транскрипции, которые, в свою очередь, непосредственно активируют гены-мишени. Второй класс генов кодирует синтез сигнальных молекул, обеспечивающих процесс межклеточной кооперации. Как минимум несколько факторов транскрипции, участвующих в процессе специализации энтодермы, продолжают экспрессироваться в тканях ЖКТ в течение всего последующего его развития, например семейства генов Fox и факторов GATA (семейство факторов транскрипции, характеризующихся способностью связываться с фрагментом дезоксирибонуклеиновой кислоты (ДНК) и с GATA-последовательностью нуклеотидов). В регуляции развития ЖКТ принимают участие, действуя в строго определенное время в различных локусах, сигнальные пути, или механизмы передачи регуляторных стимулов внутри клетки через определенную цепь молекул-посредников, представители группы факторов роста — трансформирующие факторы роста b в том числе непосредственно трансформирующий фактор роста b (TGF-b), и белок морфогенеза костной ткани (BMP), а также сигнальные пути hedgehog (сигнальные системы млекопитающих, представленные тремя различными белками). Продемонстрировано, что фактор транскрипции Sox-17 играет крайне важную роль в развитии энтодермы у мышей на ранних этапах эмбриогенеза, однако роль Sox-17 в дальнейшем не установлена. В последнее время выявлено, что сигнальный путь Notch (рецепторзависимый латентный фактор транскрипции, локализованный в клеточной цитоплазме) также выполняет важную функцию в регуляции развития ЖКТ. До настоящего времени не удалось выяснить, существует ли «главный ген», запускающий формирование энтодермы и последующее развитие ЖКТ. При исследовании некоторых животных моделей установлены гены, присутствие которых является необходимым и в то же время достаточным условием для формирования энтодермы, например ген mixer у шпорцевых лягушек. Ген мышей mixl1 играет основную роль в дифференцировке эмбриональных клеток в энтодерму. У других лабораторных животных обнаружены гены, необходимые для обеспечения данного процесса, однако недостаточные для формирования полноценной энтодермы. В исследованиях на лабораторных животных установлены начальные этапы индукции образования энтодермы. Энтодерма у мышей формируется из передней части первичной полоски эпибласта. Ключевой регулятор ранних этапов развития энтодермы — nodal-лиганд факторов роста суперсемейства TGF-b. Nodal экспрессируется в расположенном в бластодерме организационном центре или в узелке первичной бороздки эмбриона мыши. В отсутствие экспрессии nodal эмбрионы мыши не способны к гаструляции или к формированию дифинитивной энтодермы. Аналогично у эмбрионов Danio rerio в отсутствие экспрессии гомологов nodal также не образуется энтодерма. Напротив, экспрессия функционально активных рецепторов к TGF-b способствует преобразованию эмбриональных клеток в энтодерму. Реализация сигнального пути TGF осуществляется посредством стабильного комплекса промежуточных белков Smad. Необходимым для развития энтодермы является белок Smad2, и нокаут этого белка приводит к нарушению формирования энтодермы задней кишки, а разные мутации в гене, кодирующем Smad2, вызывают различные аномалии передней/головной кишки. FoxA2 также играет важную роль в развитии энтодермы, являясь мишенью сигнального пути, запущенного фактором nodal через белок Smad2. Smad2 и Smad3 совместно регулируют процесс развития энтодермы у мышей, оказывая различное влияние на формирование передней кишки и задней кишки. Кроме того, сигнальная система Smad необходима для развития печени. Основную роль в формировании печени играет ген Hex, экспрессия которого отсутствует либо существенно снижена при нарушенной продукции белков Smad. Этапы развития желудочно-кишечного тракта у плода
Установлено, что энтодерма находится в тесном контакте с мезодермой на протяжении всего ЖКТ. В экспериментальных исследованиях, выполненных на культурах тканей, было показано, что формирование энтодермы и ее дифференцировка в отдельные органы представляет собой результат взаимодействия мезодермы и энтодермы. Раньше всего было показано, что для формирования переднезадней оси энтодермы мышей необходимо сигнальное воздействие мезодермы на энтодерму посредством фактора роста фибробластов (FGF). Другие члены семейства FGF и рецепторы к ним необходимы для формирования печени. Еще три семейства генов крайне необходимы в процессе взаимодействия мезодермы и энтодермы — гены, кодирующие факторы сигнальных путей hedgehog и BMP, гены box. У почвенной нематоды Caenorhabditis elegans для специализации клеток, из которых развивается кишечный эпителий, необходимы два гена, кодирующие факторы транскрипции GATA, в то время как фактор GATA у плодовых мушек рода Drosophila кодируется одним геном serpent, наличие которого нужно для дифференцировки кишечной энтодермы. В кишечнике позвоночных экспрессируются три фактора группы GATA. Предполагается, что в процессах пролиферации и дифференцировки кишечного эпителия факторы GATA4, -5 и -6 выполняют различные функции, однако их функциональное значение на ранних этапах развития кишечной трубки у млекопитающих пока не установлено. Помимо факторов GATA, представители семейства forkhead-related факторов (кодируемые генами Fox) и сигнального пути WNT/Tcf являются важными регуляторами развития энтодермы. У позвоночных важную роль в инициации формирования энтодермы играют представители суперсемейства TGF-b. Молекула цитоскелета ELF3 задействована в сигнальном пути через факторы TGF-P, а также необходима для развития энтодермы. В отсутствие гена, кодирующего ELF3, у мышей полностью отсутствует кишечная энтодерма. Многочисленные факторы транскрипции, первоначально считавшиеся специфическими для формирования печени, играют роль и в развитии кишечника. При изучении эмбрионов мыши было отмечено, что некоторые из факторов транскрипции имеют характерные паттерны экспрессии, что позволило предположить их участие в развитии кишечника. Например, нуклеарный печеночный фактор (HNF) 3b (сейчас известный как FoxA2) оказался важным элементом в процессе самых ранних этапов дифференцировки ЖКТ и продолжает экспрессироваться в производных энтодермы у взрослых. Гомозиготные мутанты с полным отсутствием HNF3b не способны к формированию первичной полоски, предшествующей образованию кишечной трубки и ряда других структур. HNF3b участвует в образовании передней и средней, но не задней кишки. Идентифицировано большое количество факторов семейства Fox, некоторые из которых характеризуются преимущественной или специфической экспрессией в кишечнике. Один из представителей данного семейства, Foxl1, в норме экспрессируется кишечной мезодермой и представляет собой основной медиатор взаимодействия между эпителием и мезенхимой. Отсутствие указанного фактора сопровождается патологической пролиферацией эпителиальных клеток и нарушением развития кишечника. Этот фактор действует через экспрессию протеогликанов, проявляющих себя как корецепторы к компонентам сигнального пути WNT, и таким образом активирует комплекс WNT/P-катенин, регулирующий пролиферацию клеток. Группа исследователей во главе с Zaret предложила модель, в которой факторы FoxA2 и GATA открывают ДНК в клетках энтодермы, обеспечивая ее доступность для последующего связывания с факторами транскрипции, регулирующими клеточно-специфические гены. Таким образом, вероятно, при развитии кишечника многочисленные представители семейства факторов Fox играют важную роль в этом сложном процессе, который окончательно не изучен. У мышей некоторые гены, гомологичные гену caudal у дрозофил, экспрессируются исключительно в кишечнике. Cdx-1, экспрессирующийся в кишечнике взрослых организмов, также широко экспрессируется в тканях развивающегося эмбриона. Другой ген, Cdx-2, экспрессируется в висцеральной энтодерме на ранних стадиях эмбрионального развития, но впоследствии его паттерн экспрессии ограничен исключительно кишечником. Чрезмерная экспрессия Cdx-2 приводит к дальнейшей дифференцировке популяции клеток кишечника, которые в норме должны оставаться недифференцированными. В то же время эктопическая экспрессия Cdx-2 в стенке желудка приводит к формированию ткани, аналогичной кишечнику. Cdx-2 представляет собой важный фактор дифференцировки кишечника, однако до настоящего времени его роль в формировании кишечной трубки на ранних стадиях эмбрионального развития точно не установлена.
– Также рекомендуем “Генетическая регуляция формирования кишечной трубки” Оглавление темы “Эмбриогенез желудочно-кишечного тракта”:
|
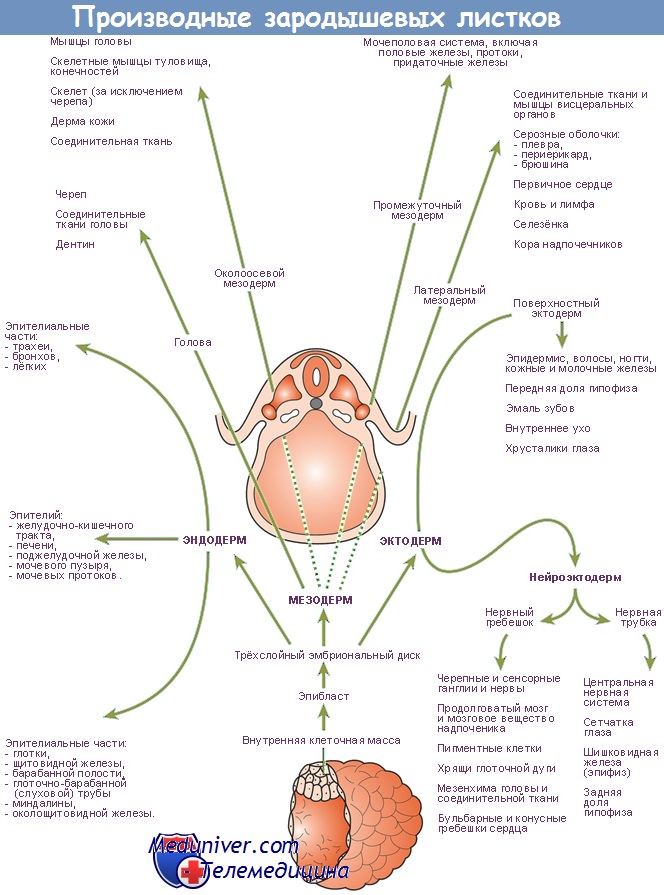
Источник
Дифференцировка энтодермы приводит к образованию в теле зародыша энтодермы кишечной трубки и формированию внезародышевой энтодермы, формирующей выстилку желточного пузырька и аллантоиса. [4]
Выделение кишечной трубки начинается с момента появления туловищной складки. Последняя, углубляясь, отделяет кишечную энтодерму будущей кишки от внезародышевой энтодермы желточного пузырька. В задней части зародыша в состав образующейся кишки входит и тот участок энтодермы, из которого возникает энтодермальный вырост аллантоиса.[5]
Из энтодермы кишечной трубки развивается однослойный покровный эпителий желудка, кишечника и их желез. Кроме того, из энтодермы развиваются эпителиальные структуры печени и поджелудочной железы.[1]
Внезародышевая энтодерма дает начало эпителию желточного мешка и аллантоиса.
Этот процесс начинается на 3-й неделе эмбриогенеза. Дорсальные участки мезодермы разделяются на плотные сегменты, лежащие по сторонам от хорды, – сомиты. Процесс сегментации дорсальной мезодермы и образования сомитов начинается в головной части зародыша и быстро распространяется в каудальном направлении.[7]
У эмбриона на 22-е сутки развития имеется 7 пар сегментов, на 25-е – 14, на 30-е – 30 и на 35-е – 43 -44 пары.В отличие от сомитов вентральные отделы мезодермы (спланхнотом) не сегментируются, а расщепляются на два листка – висцеральный и париетальный. Небольшой участок мезодермы, связывающий сомиты со спланхнотомом, разделяется на сегменты – сегментные ножки (нефрогонотом). На заднем конце зародыша сегментации этих отделов не происходит. Здесь взамен сегментных ножек располагается несегментированный нефрогенный зачаток (нефрогенный тяж). Из мезодермы зародыша развивается также парамезонефральный канал.[3]
Сомиты дифференцируются на три части: миотом, дающий начало поперечнополосатой скелетной мышечной ткани, склеротом, являющийся источником развития костных и хрящевых тканей, а также дерматом, формирующий соединительнотканную основу кожи – дерму.[6]
Из сегментных ножек (нефрогонотомов) развиваются эпителий почек, гонад и семявыводящих путей, а из парамезонефрального канала – эпителий матки, маточных труб (яйцеводов) и эпителий первичной выстилки влагалища.
Париетальный и висцеральный листки спланхнотома образуют эпителиальную выстилку серозных оболочек – мезотелий. Из части висцерального листка мезодермы (миоэпикардиальная пластинка) развиваются средняя и наружная оболочки сердца – миокард и эпикард, а также корковое вещество надпочечников.[1.2]
Мезенхима в теле зародыша является источником формирования многих структур – клеток крови и кроветворных органов, соединительной ткани, сосудов, гладкой мышечной ткани, микроглии.[2] Из внезародышевой мезодермы развивается мезенхима, дающая начало соединительной ткани внезародышевых органов, – амниона, аллантоиса, хориона, желточного пузырька.[7]
Соединительная ткань эмбриона и его провизорных органов характеризуется высокой гидрофильностью межклеточного вещества, богатством гликозаминогликанов в аморфном веществе. Соединительная ткань провизорных органов дифференцируется быстрее, чем в органных зачатках, что обусловлено потребностью в установлении связи зародыша с материнским организмом и обеспечении их развития (например, плацента). [5] Дифференцировка мезенхимы хориона наступает рано, но происходит не одновременно по всей поверхности. Наиболее активно процесс идет в области развития плаценты. Здесь же появляются и первые волокнистые структуры, которые играют важную роль в формировании и укреплении плаценты в матке. При развитии волокнистых структур стромы ворсин последовательно образуются сначала аргирофильные преколлагеновые волокна, а затем коллагеновые.[3]
На 2-м месяце развития в зародыше человека раньше всего начинается дифференцировка скелетогенной и кожной мезенхимы, а также мезенхимы стенки сердца и крупных кровеносных сосудов.
Артерии мышечного и эластического типа эмбрионов человека, а также артерии стволовых (якорных) ворсин плаценты и их разветвлений содержат десминотрицательные гладкие миоциты, обладающие свойством более быстрого сокращения.[1.1]
На 7-й неделе развития зародыша человека в кожной мезенхиме и мезенхиме внутренних органов появляются мелкие липидные включения, а позднее (8-9-я неделя) происходит формирование жировых клеток. Вслед за развитием соединительной ткани сердечно-сосудистой системы дифференцируется соединительная ткань легких и пищеварительной трубки. [6] Дифференцировка мезенхимы у зародышей человека (длиной 11-12 мм) на 2-м месяце развития начинается с увеличения количества гликогена в клетках. В этих же участках возрастает активность фосфатаз, а в дальнейшем в ходе дифференцировки накапливаются гликопротеины, синтезируются РНК и белок.
Плодный период. Плодный период начинается с 9-й недели и характеризуется значительными морфогенетическими процессами, протекающими в организме как плода, так и матери.
Источник
Первичная эктодерма образует: нервную трубку, ганглиозные пластинки, плакоды. Кожную эктодерму, прехордальную пластинку и внезародышевую эктодерму (эпителий амниона).
Дифференцировка начинается с процесса нейруляции – нервная пластинка углубляется в виде желобка, а затем замыкается в трубку. Замыкание начинается в шейном отделе и распространяется назад. В головном отделе процесс задерживается, т.к. нужно создать достаточно клеток для зачатка головного мозга.
При смыкании трубки из краев нервного желобка выселяются клетки, которые ложатся по обе стороны от трубки. Так образуется нейральный гребень. Его клетки способны к миграциям. Отсюда клетки мигрируют двумя потоками. Один идет наверх, внедряясь в наружные покровы тела. Это будущие меланоциты кожи. Другой поток идет в брюшном направлении, формируя симпатические и парасимпатиченские ганглии, а также мозговое вещество надпочечников. Остатки гребня сегментируются и дают начало спинномозговым узлам.
После нейруляции в наружном листке остаются островками нервного зачатка, называемые «плакоды». Они участвуют в формировании органов чувств. После выселения нервной трубки говорят о вторичной или кожной эктодерме. Из нее развивается эпидермис со своими производными, а также многослойные неороговевающие эпителии, которые служат источниками для целого ряда структур (см. слайд).
Вопрос 5
Дифференцировка мезодермы.
Мезодерма подвергается дифференцировке начиная с 20-х суток эмбриогенеза. Дифференцируется следующим образом. Вначале она представляет собой более или менее рыхлое скопление клеток (пресомитная мезодерма), а затем разделяется на дорзальную и вентральную мезодерму. Дорзальная мезодерма по длине зародыша разделяется на сегменты — сомиты. Сегментация дорзалыюй мезодермы начинается на переднем конце и быстро распространяется в каудальном направлении. Количество сомитов нарастает во времени; на 22-е сутки их 7 пар, 25-е — 14, 30-е — 30, 35-е — 43—44 пары. Образование сомитов настолько важный этап эмбриогенеза, что его часто выделяют как отдельный сомитный период.
Каждый сомит, в свою очередь, дифференцируется на 3 эмбриональных зачатка: наружный — дерматом, средний — миотом, внутренний — склеротом. Из дерматома в дальнейшем сформируется дерма кожи. Миотом послужит источником скелетной поперечнополосатой мышечной ткани. Из склеротома образуются костные и хрящевые ткани.
Вентральная мезодерма (спланхнотом) не подвергается сегментации. Она разделяется на два листка — висцеральный и париентальный листки спланхнотома. Между ними находится вторичная полость тела — целом. Из листков спланхнотома развиваются: мезотелий серозных оболочек, поперечнополосатая сердечная мышечная ткань, корковое вещество надпочечников, эпителий гонад.
Между дорзальной и вентральной мезодермой находится промежуточная мезодерма, или нефротом. В передних отделах тела зародыша он сегментируется на сомитные ножки, в задних же сегментации не подвергается. Из сегментированных отделов нефротома последовательно развиваются предпочка и первичная почка, а в мужском организме — и выносящие канальцы придатка яичка. Несегментированная часть нефротома называется нефрогенной тканью. Она служит источником для формирования окончательной почки.
Вопрос 6
Одновременно с дифференцировкой компактных зачатков, все промежутки между ними заполняются отростчатыми, рыхло лежащими клетками. Их объединяют понятием «мезанхима». Мезенхима – это очень гетерогенная популяция дифференцирующихся клеток, мигрирующих из состава всех листков (можно говорить о нейромезенхиме, энтомезенхиме и т.д.) и многих зачатков (например из миотома – в формирующуюся конечность). Мезодермальная мезенхима также формирует ряд т.н. диффузных зачатков. Это ангиодермальный зачаток, из которого формируется эндотелий кровеносных сосудов и клетки крови, гладкомышечный зачаток, зачаток волокнистых соединительных тканей.
Дифференцировка мезенхимы происходит под индуцирующим влиянием со стороны соседних эмбриональных структур. Например, энтодерма инициирует дифференцировку мезенхимных клеток в эндотелий сосудов.
Вопрос 7
Дифференцировка энтодермы
С момента появления туловищной складки выделяется кишечная энтодерма. По мере углубления складки она все больше обособляется от внезародышевой энтодермы, выстилающей желточный мешок. Кишечная трубка является источником для образования эпителия желудка, кишечника, печени, желчного пузыря и поджелудочной железы. Все энтодермальные эпителии однослойны. Мезенхима вокруг первичной кишки преобразуется в соединительную ткань и гладкую мускулатуру.
От задней части кишки в амниотическую ножку врастает энтодермальный эпителий аллонтоиса.
На 4-й неделе в эктодерме образуется впячивание, которое прорываетсяв кишку. Так возникает ротовое отверстие. Немного позже в состав головной части кишечной трубки включится материал прехордальной пластинки.
Вопрос 8
Итак к концу первого месяца завершается нотогенез.
Нотогенез — это процесс образования осевого комплекса зачатков. Осевым он называется потому, что образовавшиеся из зародышевых листков эмбриональные зачатки располагаются по длинной, сагиттальной оси тела. В основе нотогенеза лежат три важных тесно взаимосвязанных процесса, происходящие в основном в течение 4-й недели эмбриогенеза.
1. Нейруляция; 2. Дифференцировка зародышевых листков, в первую очередь мезодермы. 3. Образование туловищных складок с отделением зародыша от внезародышевых органов и образованием кишечной трубки. Осевой комплекс включает следующих зачатки :
1. Кожная эктодерма.
2. Нервная трубка и ганглиозные пластинки.
3. Сомиты, состоящие из дерматома, миотома и склеротома.
4. Нефротом.
5. Спланхнотом.
6. Хордальный отросток
7. Кишечная трубка.
8. Мезенхима.
К 9-й неделе развития (начало 3-го месяца) длина зародыша составляет 40 мм, а масса около 5 г. и он имеет закладки всех систем, а его сердце уже начинает сокращаться. Однако период гисто- и органогенеза будет продолжаться вплоть до рождения.
Вопрос 9.
Механизмы эмбриогенеза.
Источником развития тканей являются эмбриональные зачатки. В свою очередь, эмбриональные зачатки развиваются из зародышевых листков в процессе их дифференцировки. В результате формируется осевой комплекс зачатков. Процесс образования тканей в эмбриогенезе из тканевых зачатков называется эмбриональным гистогенезом. Механизмы гистогенеза достаточно сложны и включают следующие компоненты:
1, Деление клеток. В результате деления клеток зачатка происходит нарастание клеточного материала, объема зачатка, достижение им критической массы, что запускает дальнейшие гистогенетические процессы. Основным видом деления клеток в ходе гистогенеза является митоз. Он может быть стволовым, асимметричным и дифференцирующим, или кван-тальным. При стволовом митозе из одной материнской стволовой клетки образуются две дочерние стволовые клетки. Для асимметричного митоза характерно то, что из двух дочерних клеток одна является стволовой, а вторая вступает на путь дифференцировки. При квантальном митозе обе дочерние клетки отличаются от стволовых, поскольку уже приступили к
дифференцировке.
2. Рост клеток. Наряду с митозом рост клеток приводит к увеличению
общей массы зачатка ткани. В его основе лежат гипертрофия и гиперпла
зия клеточных органелл, накопление включений.
3. Гибель клеток.
Запрограммированная гибель клеток, или апоптоз. По своему значе-
нию клеточная гибель не менее важна для гистогенетических процессов,чем деление клеток. В результате апоптоза регулируется число клеток в развивающейся ткани, происходит ее перестройка, исчезают рудиментарные зачатки, элиминируются мутировавшие и дефектные клетки. Любопытно, что в некоторых случаях в ходе гистогенеза сразу образуется заведомо больше клеток, чем их необходимо для развития ткани, и это создает определенный материальный базис гистогенеза. В последующем лишние клетки погибают, причем уничтожаются менее полноценные или дефектные клетки. Особенно это явление выражено в нервной ткани, где в ходе гистогенеза гибнет от 50 до 85 % всех нейронов.
4. Миграция клеток. Различают пассивную и активную миграцию кле
ток. Пассивная миграция — миграция в результате давления соседних
клеток. Активная миграция клеток происходит за счет работы внутрикле
точных сократительных структур, связанных через подмембраниый слой с
поверхностными рецепторами.
5. Адгезия клеток и межклеточные взаимодействия (механизмы кле
точной адгезии — см. цитологию). Для образования ткани необходимо,
чтобы клетки зачатка совершили миграционные процессы, а затем сфор
мировали клеточные ансамбли. Инициация миграции связана с потерей
клетками зачатка адгезионных молекул (эта ситуация определяется как
конец адгезии — начало миграции). После начала миграции клеточная ад
гезия контролирует миграцию клеток: мигрирующие в ходе гистогенеза
клетки узнают на поверхности других клеток или во внеклеточном мат-
риксе адгезионные молекулы, что обеспечивает целенаправленность мигра
ции. После завершения миграции начинается процесс формирования нуж
ных клеточных ансамблей. При этом в завершивших миграцию клетках
вновь появляются молекулы адгезии, и между клетками устанавливаются
взаимодействия (конец миграции — начало адгезии).
6. Детерминация (процесс определения пути, программы развития эмб
риональных зачатков в направлении той или иной дефинитивной ткани).
Механизм детерминации связан со стойкой репрессией одних и дерепрес-
сией других генов, необходимых для развития клеток будущей ткани в
нужном направлении.
7. Дифференцировка — стойкое структурно-функциональное измене
ние ранее однородных клеток, приобретение ими специфических черт стро
ения для выполнения специфических функций. Молекулярно-генетических
основы дифференцировки — транскрипция, сплайсинг РНК, ее процессинг,
трансляция, т.е. синтез специфических и-РНК и на них — специфических
белков. Морфологической основой дифференцировки является образование
из специфических белков специфических клеточных органелл.
8. Эмбриональная индукция. Эмбриональная индукция — это на
правление гистогенетических процессов в нужное русло путем выделения
одним зачатком веществ — индукторов, действующих на другой зачаток. В
качестве эмбриональных индукторов могут выступать не только химические индукторы, биологически активные вещества и гормоны (вторичные индукторы), но и самые обычные факторы: питательные вещества, уровень рН, концентрация электролитов, кислорода и др. (первичные индукторы).
Органогенез — процесс образования органов и систем органов из эмбриональных зачатков. Этот процесс протекает обычно параллельно с гистогенезом, т.е. с образованием тканей в составе будущих органов, и отделить два процесса друг от друга невозможно. В процессе органогенеза организм зародыша разделяется на относительно независимо развивающиеся местные системы, дающие орган. Многие механизмы гистогенеза и органогенеза являются общими.
На определенном этапе органогенеза образование органов становится невозможным без участия регуляторных механизмов. Таковыми являются нервные, эндокринные и иммунные регуляторные влияния.
Дата добавления: 2015-11-05; просмотров: 4098 | Нарушение авторских прав | Изречения для студентов
Читайте также:
Рекомендуемый контект:
Поиск на сайте:
© 2015-2021 lektsii.org – Контакты – Последнее добавление
Источник