Железы желудочно кишечного тракта

Железы пищеварительного тракта. Строение желез пищеварительного тракта
На всем протяжении пищеварительного тракта секреторные железы выполняют две ведущие функции: (1) секрецию пищеварительных ферментов, которая начинается в полости рта и заканчивается в дистальном отделе подвздошной кишки; (2) выработку слизи слизистыми железами на всем протяжении от рта до ануса для смазки и защиты всех участков пищеварительного тракта.
Основная часть пищеварительных секретов вырабатывается только в ответ на присутствие пищи в пищеварительном тракте, а количество секрета в каждом отделе тракта почти всегда соответствует количеству, необходимому для оптимального переваривания пищи. К тому же в некоторых отделах гастроинтестинального тракта даже состав ферментов и другие параметры секреции изменяются в соответствии с находящимся там типом пищи.
Секрецию в пищеварительном тракте обеспечивают несколько типов желез. Во-первых, па поверхности эпителия большей части желудочно-кишечного тракта присутствуют миллиарды одноклеточных слизистых желез, называемых простыми слизистыми клетками, или бокаловидными клетками (из-за их внешнего вида). Эти клетки функционируют главным образом в ответ на местное раздражение эпителия: они выталкивают слизь прямо на эпителиальную поверхность. Слизь действует как смазка, защищающая поверхность от ссадин и переваривания.
Во-вторых, большая часть поверхности желудочно-кишечного тракта содержит углубления, которые представляют собой инвагинации эпителия в подслизисгый слой. В тонкой кишке эти углубления, названные криптами Либсркюна, — глубокие и содержат специализированные секретирующие клетки..
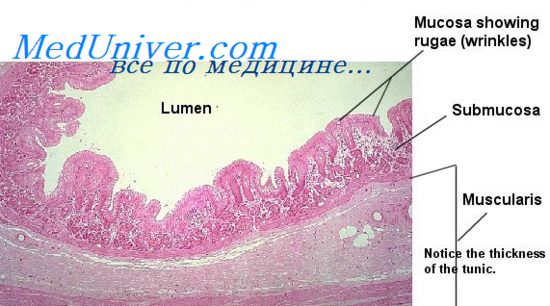
В-третьих, в желудке и верхнем отделе двенадцатиперстной кишки находится большое количество глубоких трубчатых желез. Типичную трубчатую железу можно увидеть на рисунке, на котором изображена кислото- и пепсиноген-секретирующая железа желудка (кислотопродуцирующая железа).
В-четвертых, есть несколько сложных желез, также связанных с желудочно-кишечным трактом, — слюнные железы, поджелудочная железа и пс чень, которые вырабатывают секреты для переваривания или эмульгирования пищи. Слюнные железы и поджелудочная железа состоят из смешанных альвеолярных желез. Эти железы лежат за пределами пищеварительного тракта, чем отличаются от всех других пищеварительных желез. Они содержат миллионы ацинусову выстланных секретирующими железистыми клетками. Ацинусы снабжают систему протоков, которые в итоге самостоятельно открываются в пищеварительный тракт.
Механическое раздражение пищей в определенном отделе желудочно-кишечного тракта обычно вызывает адекватную секрецию желез этой области и часто — смежных с ней областей. Часть этих местных эффектов, особенно секреция слизи слизистыми клетками, происходит за счет непосредственного раздражения пищей поверхности железистых клеток.
Кроме того, местная стимуляция эпителия активирует энтеральную нервную систему кишечной стенки. Раздражителями могут служить следующие стимулы: (1) тактильная стимуляция; (2) химическое раздражение; (3) растяжение кишечной стенки. В результате стимуляции как слизистые клетки эпителиальной поверхности кишки, так и глубокие железы кишечной стенки рефлекторно усиливают свою секрецию.
– Вернуться в оглавление раздела “Физиология человека.”
Оглавление темы “Двигательная активность желудочно-кишечного тракта”:
1. Регуляция глоточной фазы глотания. Пищеводная фаза глотания
2. Расслабление желудка при глотании. Функция пищеводного сфинктера при глотании
3. Двигательная активность желудка. Формирование химуса в желудке
4. Опорожнение желудка. Регуляция опорожнения желудка и факторы влияющие на нее
5. Факторы торможения опорожнения кишечника. Роль жиров в торможении эвакуации из желудка
6. Перистальтика тонкого кишечника. Виды двигательной активности тонкого кишечника
7. Усиленная перистальтика тонкого кишечника. Физиология илеоцекального клапана
8. Перистальтика толстого кишечника. Формы двигательной активности толстого кишечника
9. Акт дефекации. Регуляция акта дефекации
10. Железы пищеварительного тракта. Строение желез пищеварительного тракта
Источник
Одноклеточные железы представлены в ЖКТ бокаловидными клетками — разновидностью эпителиальных клеток, специализирующихся на секреции слизи (рис. 5.7, а).
Бокаловидные клетки в больших количествах располагаются среди обычных эпителиальных клеток во всех отделах пищеварительной системы и присутствуют в структурах многоклеточных желез. Секретом бокаловидных клеток является слизь, образованная муцинами, водой и солями.
Муцины являются гликопротеинами, на углеводную часть которых приходится 60—80% молекулярной массы. Слизь обволакивает изнутри всю эпителиальную выстилку пищеварительной трубки, при этом несколько отличаясь по составу в разных ее отделах. Функции слизи разнообразны, но могут быть сведены к смазке и защите эпителия. Выполняя роль смазки, слизь облегчает продвижение пищевой массы, снижая ее трение о стенки, а также участвует в формировании каловых масс. Слизь обеспечивает механическую защиту эпителия кишечника, предотвращая его непосредственное соприкосновение с пищевыми частицами. Благодаря хорошим адгезивным свойствам слизь защищает эпителий от действия ферментов пищеварительных соков, а благодаря амфотерным свойствам — от резких перепадов pH. Помимо этого, слизь способна выполнять функции «бактериальной ловушки», осуществляя антибактериальную защиту пищеварительной системы. Следующим по анатомической сложности типом желез являются секреторные крипты.
Рис. 5.7. Различные анатомические типы желез пищеварительной системы:
а — одноклеточная железа (бокаловидная клетка); б — секреторная крипта; в — простая трубчатая железа; г, д — усложненные трубчатые железы;
е — сложная железа
Секреторные крипты представляют собой впячивания эпителия в подслизистый слой стенки пищеварительной трубки. Крипты присутствуют в стенках тонкого и толстого кишечника (рис. 5.7, б). В тонком кишечнике крипты располагаются между кишечными ворсинками (либеркюновы крипты). На 1 мм2 стенки кишки насчитывается от 80 до 100 крипт, имеющих диаметр около 0,07 мм и длину от 0,25 до 0,5 мм. В стенках крипты лежат клетки, секретирующие ферменты, клетки, секретирующие слизь, и клетки, секретирующие воду и электролиты. На дне каждой крипты располагаются активно делящиеся клетки, за счет которых происходит обновление всего эпителия. Новообразованные клетки постепенно мигрируют к вершинам ворсинок, где происходит их слущивание.
Простые трубчатые железы присутствуют в желудке и в верхнем отделе двенадцатиперстной кишки (рис. 5.7, в — д). Они берут начало в глубине слизистой оболочки и характеризуются трубчатой формой (но могут быть и разветвления) и отсутствием протоков. В стенках трубчатых желез лежат секреторные клетки разных типов, что позволяет вырабатывать сложные многокомпонентные секреты. В разных отделах желудка трубчатые железы вырабатывают секрет разного состава. Всего в желудке насчитывается около 40 млн трубчатых желез.
Сложные железы являются высокоспециализированными секреторными органами, для которых, помимо секреторных конечных участков, характерно наличие системы протоков (рис. 5.7, е). Секреторные конечные участки (ацинусы) производят первичный секрет, который затем преобразуется под влиянием эпителиальных клеток стенок протоков. К сложным железам относятся крупные слюнные железы, железы пищевода, железы Бруннера в двенадцатиперстной кишке, поджелудочная железа и печень.
Механизм секреции органических веществ принципиально сходен в секреторных клетках всех пищеварительных желез. Вместе с кровью по капиллярам, подходящим к базальной части секреторных клеток, поступают исходные вещества, необходимые для последующего синтеза. Многочисленные митохондрии, сконцентрированные в базальной части клеток, синтезируют ЛТФ. ЛТФ расходуется в процессе синтеза органических веществ в ЭПР и комплексе Гольджи. Белки синтезируются на рибосомах в шероховатом ЭПР. По системе цитоскелета синтезированные вещества транспортируются в цистерны комплекса Гольджи, где концентрируются и модифицируются, после чего внутри секреторных везикул выделяются в цитоплазму. Секреторные везикулы скапливаются в апикальной части клетки и при поступлении соответствующего нервного или гормонального сигнала экзоцитируют. Таким образом, произведенный секрет оказывается выведенным из клетки (рис. 5.8).
Обязательными компонентами секрета пищеварительных желез, помимо органических веществ, являются вода и электролиты.
Механизм секреции воды и электролитов запускается ВНС или под воздействием гормонов. Мишенью для нервных или гормональных стимулов является базальная часть секреторной клетки, к которой подходят нервные окончания и кровеносные капилляры. Под влиянием нервных или гормональных стимулов активируется котранспортер, доставляющий в цитоплазму катионы К+, Na+ и два аниона С1~ за один цикл своей работы. Избыток ионов Na+
выводится из клетки за счет работы Ыа+/К+-АТФ-азы, а избыток К * — через калиевые каналы. Накопившиеся в цитоплазме анионы С1_ выводятся в просвет железы, а вслед за ними по межклеточным пространствам следуют ионы Na+. Выход ионов Na+ и С1_ в просвет железы создает осмотический градиент между кровью, тканевой жидкостью и заполняющим просвет секретом. За счет этого градиента в просвет железы поступает вода (рис. 5.9). Некоторое количество воды может проходить через клетку (трансцеллюлярно), также используя осмотический градиент.
Рис. 5.8. Механизм секреции органических веществ
Рис. 5.9. Механизм секреции NaCl и воды секреторными клетками желез
Источник
А. Большие слюнные железы (рис. 12-23,12-50,12-51). К ним относят три пары слюнных желёз: околоушные, подчелюстные и подъязычные. Это сложные трубчато-альвеолярные железы. В зависимости от характера секрета, различают белковые, слизистые и смешанные концевые отделы. Слюнные железы, содержащие в концевых отделах преимущественно белковые или слизистые клетки, относят соответственно к белковым или слизистым железам. Смешанные железы в концевых отделах содержат как белковые, так и слизистые клетки. Околоушная железа — чисто белковая, подъязычная железа — преимущественно слизистая и подчелюстная железа — смешанная. Секрет всех слюнных желёз образует слюну в количестве около I л в сутки. Слюна гипотонична по отношению к плазме. Она смачивает и очищает полость рта, Присутствующие в слюне лизоцим, лактоферрин и IgA контролируют бактериальную флору полости рта. Амилаза слюны расщепляет остатки крахмала вокруг зубов.
- Секреторный отдел. В базальной части клеток секреторного отдела находятся ядро и гранулярная эндоплазматическая сеть, наиболее развитая в белковых клетках. Как в слизистых, так и в белковых клетках секреторные гранулы скапливаются в апикальной части. Секреторные гранулы белковых клеток содержат амилазу и гликопротеины. Секреторные гранулы слизистых клеток крупнее, чем в белковых, содержат муцин и гликопротеины. Периферическую часть секреторных отделов занимают миоэпителиальные клетки.
- Выводной проток. От концевых отделов начинается разветвлённая система выводных протоков: вставочный отдел, исчерченный проток (слюнная трубка), внутридольковый и междольковый протоки.
а. Вставочный отдел выстлан плоским или кубическим эпителием.
б. Исчерченный проток представлен цилиндрическими эпителиальными клетками (рис. 12-24), образующими в базальной части многочисленные инвагинации, значительно увеличивающие площадь клеточной мембраны для транспорта ионов. Здесь расположены многочисленные митохондрии удлинённой формы, ориентированные параллельно апикально-базальной оси клетки. Эпителиальные клетки исчерченного протока переводят изотонический секрет, образующийся в концевых отделах, в гипотонический окончательный секрет, входящий в состав слюны (рис. 12-25).
- Нервный контроль секреции. Парасимпатические холинергические волокна заканчиваются на клетках секреторного отдела и выводных протоков и значительно усиливают секреторную активность железы.
Б. Поджелудочная железа состоит из экзокринной и эндокринной частей. Эндокринная часть (островки Лангерханса) рассмотрена в IV I. Экзокринная часть участвует в переваривании белков, липидов и углеводов. Секретируемый железой бикарбонат вместе с бикарбонатом двенадцатиперстной кишки и гепато-билиарной системы участвует в нейтрализации соляной кислоты, поступающей из желудка в двенадцатиперстную кишку.
- Структура экзокринной части (рис. 12-26, 12-27 и 12-52). В железе различают дольки, состоящие из ацинусов и начальных отделов выводных протоков. Взаимоотношения между ними представлены на рис. 12-26 и 12-27. Протоки выводят секреторные продукты ацинуса и выделяют бикарбонат. В центре ацинусов расположены т.н. центроацинозные клетки (рис. 12-27). От них начинаются выводные протоки. Кубический или цилиндрический эпителий внутридольковых выводных протоков переходит в цилиндрический эпителий междольковых протоков. Среди эпителиальных клеток присутствуют энтероэндокринные клетки.
Ацинозные клетки синтезируют, хранят и секретируют пищеварительные ферменты.
- Структура (рис. 12-28). Секреторные клетки характеризуются выраженной полярной дифференцировкой. Апикальная их часть содержит многочисленные зимогенные гранулы с пищеварительными ферментами. Ядро смещено в базальную часть, где присутствуют хорошо развитая гранулярная эндоплазматическая сеть,

Рис. 12-23. Подчелюстная слюнная железа содержит белковые и белково-слизистые (смешанные) секреторные отделы. Секреторные отделы переходят во вставочный отдел. По мере продвижения секрета по выводным протокам увеличивается количество слоев клеток в эпителии [из Braus из Bloom W, FawceH DW, 1968]

Рис. 12-24. Эпителиальная клетка исчерченного протока большой слюнной железы. В базальной части клетки содержатся митохондрии, имеются многочисленные инвагинации плазматической мембраны. Крупное округлое ядро занимает центральную область клетки. Апикальная часть заполнена везикулами. Комплекс Гольджи расположен над ядром [из Lentz TL, 1971]
Рис. 12-25. Транспорт ионов и глюкозы в околоушной слюнной железе. Секреторные отделы содержат изотонический по сравнению с плазмой секрет. Эпителиальные клетки протоков активно выкачивают Na* и Cl” из жидкости в просвете протока и секретируют в неё Kt и глюкозу. В результате образуется гипотонический (по сравнению с плазмой) окончательный секрет [из Davenport HW, 1977]
 Рис. 12-26. Организация ацинусов и внутридольковых протоков в поджелудочной железе. Состоящие из секреторных клеток ацинусы переходят в короткие вставочные протоки, начинающиеся от центроацинозных клеток. Далее секрет поступает во внутридольковые, а затем в междольковые протоки. На рисунке показаны различные варианты взаимоотношений между ацину- сами и внутридольковыми протоками [из Akao et al, 1977]
Рис. 12-26. Организация ацинусов и внутридольковых протоков в поджелудочной железе. Состоящие из секреторных клеток ацинусы переходят в короткие вставочные протоки, начинающиеся от центроацинозных клеток. Далее секрет поступает во внутридольковые, а затем в междольковые протоки. На рисунке показаны различные варианты взаимоотношений между ацину- сами и внутридольковыми протоками [из Akao et al, 1977]
Рис. 12-28. Секреторная клетка в аци- нусе поджелудочной железы —
классический пример полярно дифференцированной эпителиальной клетки. Круглое ядро с выраженным ядрышком находится в базальной части. Практически весь объём цитоплазмы этой части клетки занимают протяжённые цистерны гранулярной эндоплазматической сети, что свидетельствует об интенсивном синтезе белка. Апикальная часть клетки заполнена крупными зимогенными гранулами, содержимое которых выделяется в просвет ацинуса. Комплекс Гольджи находится между ядром и скоплением зимогенных гранул [из Lentz Т, 1971]
свободные рибосомы и митохондрии. Область между зимогенными гранулами и ядром занимает комплекс Гольджи. В цитоплазме апикальной части найдены акти- новые микрофиламенты, образующие сеть, и микротрубочки, участвующие во внутриклеточном транспорте зимогенных гранул и освобождении их содержимого во внеклеточное пространство.
- Межклеточные контакты. Мембраны соседних ацинозных клеток в апикальной части соединены при помощи плотных контактов, промежуточных контактов и десмосом. Все вместе эти контакты образуют соединительный комплекс, служащий барьером для крупных молекул, но проницаемым для воды и ионов. Кроме того, ацинозные клетки связаны при помощи щелевых контактов, обеспечивающих электрическое сопряжение и передачу ионов и низкомолекулярных веществ от клетки к клетке.
- Регуляция. Ацетилхолин (через м-холинорецепторы) и нейропептиды усиливают секреторную активность ацинозных клеток (рис. 12-29). Симпатические нервные волокна через адренорецепторный вход тормозят секреторную функцию ацинозных клеток.
- Функция. В поджелудочной железе вырабатываются панкреатический сок и ферменты.
а. Панкреатический со к (1,5-2 л в сутки) изотоничен плазме крови, имеет pH — 8-8,5 из-за высокого содержания бикарбоната, который нейтрализует кислую реакцию химуса (пищевые массы, смешанные с желудочным соком).
б. Ферменты поджелудочной железы играют ключевую роль в переваривании белков, жиров и углеводов. Оптимум действия ферментов поджелудочной железы приходится на pH — 7-8. Ферменты вырабатываются в виде предшественников, которые активируются в просвете кишки.
- Протеазы — ферменты, расщепляющие белки (трипсин, химотрипсин, карбокси- пептидазы А и В, эластаза). Трипсиноген (предшественник трипсина) активируется энтерокиназой, вырабатываемой в двенадцатиперстной кишке. Трипсин катализирует собственную активацию, активацию химотрипсиногена (предшественник химотоипсина) и поокаэбоксипептилаз.

Рис. 12-29. Рецепторные входы н внутриклеточные механизмы усиления секреторной активности ацинозных клеток поджелудочной железы. AX — ацетилхолин, P — вещество P [из Pandol SJ, Isenberg JI, 1990)
- Ферменты, расщепляющие жиры. Панкреатическая липаза, фосфолипазы Al, А2, лецитиназа.
- а-Амилаза — панкреатический фермент, расщепляющий углеводы.
- Нуклеазы — ферменты, расщепляющие нуклеиновые кислоты (ДНКаза, РНКаза).
В. Печень (рис. 12-30, 12-53). Разобраться в структуре печени несложно, если принять во
внимание следующие обстоятельства.
Кровь в орган поступает по двум сосудам — артериальная по a. hepatica (20-30% получаемой печенью крови) и венозная по v. porta (70-80% получаемой печенью крови), а оттекает по одному (v. hepatica).
Гепатоциты омываются смешанной кровью, находящейся в синусоидах.
Гепатоциты можно рассматривать как клетки с внешней секрецией и одновременно как клетки с внутренней секрецией. При этом не имеется в виду, что гепатоциты — эндокринные клетки, хотя они секретируют в кровь разные биологически активные вещества; имеется в виду, что гепатоциты, как и эндокринные клетки, характеризуются интимными отношениями с кровеносным руслом: для гепатоцитов характерен мощный обмен разными веществами с кровью — как секреция в кровь, так и поглощение из крови.
- Морфофункциональные единицы печени — классическая и портальная дольки, а также ацинус.
а. Классическая долька (рис. 12-31). Эта морфофункциональная единица имеет гексагональную форму. В центре расположена центральная вена, к которой сходятся печёночные тяжи, состоящие из гепатоцитов. Между тяжами залегают синусоиды. В области стыков нескольких классических долек расположена портальная зона.
- Портальная зона (т.н. триада). Междольковые сосуды: междольковые артерия, вена, жёлчный проток и лимфатический междольковый сосуд; кровь в синусоиды поступает из междольковых артерии и вены (бассейн воротной вены), а собирается в один коллектор (бассейн нижней полой вены, начинающийся от центральной вены).
- Синусоиды печени — анастомозирующие пустоты между анастомозирующими тяжами гепатоцитов. В синусоидах печени находится смешанная кровь.
(а) Кровоток. Кровь в классическую дольку поступает из междольковой артерии (обогащённая O2) и междольковой вены (богатая питательными веществами) соответственно по терминальным печёночным артериолам и терминальным воротным венулам. Эти сосуды открываются в синусоиды, по которым смешанная кровь направляется к центральной (терминальной печёночной) вене и далее по печёночным венам попадает в нижнюю полую вену.
(б) Пространство Дйссе — пространство между гепатоцитами и эндотелиальными клетками синусоидов. В пространство обращены микроворсинки гепатоцитов. Здесь расположены ретикулиновые волокна, поддерживающие структуру синусоидов; встречаются жиронакапливающие клетки.
б. Портальная долька — структура треугольной формы. Портальная зона образует её центр, а центральные вены трёх смежных классических долек — вершины.
в. Ацинус — структурно-метаболическая единица печени, имеющая форму ромба, вершины которого образованы центральными венами соседних гексагональных печёночных долек и смежными портальными зонами. Часть ацинуса, расположенная вблизи сосудов, кровоснабжается лучше других его отделов (зона I на рис. 12-31). Наружная же часть ацинуса, локализованная вблизи центральных вен (зона 3 на рис. 12-31), получает менее оксигенированную кровь. Поэтому структуры этой зоны ацинуса более уязвимы при интоксикациях и дефиците питательных веществ.

Рнс. 12-30. Строение печени. На переднем плане схемы представлены компоненты портальной зоны: междольковые артерия, вена и жёлчный проток. Кровь из сосудов портальной зоны поступает в синусоиды, радиально сходящиеся к центральной вене. Паренхиму печени образуют тяжи гепатоци- тов. Они формируют жёлчные капилляры, из которых жёлчь поступает в междольковые жёлчные протоки. Синусоиды выстланы эндотелиальными клетками, между которыми встречаются клетки фон Купффера [из Junqueira LC, Cameiro J, 1991]
- Желчевыводящие пути. Жёлчные капилляры (гепатоциты) -gt; холангиолы —gt; мелкие жёлчные протоки -gt; междольковые жёлчные протоки (кубический эпителий) -> крупные септальные и трабекулярные протоки (цилиндрический эпителий) -gt; внутрипечёночные протоки —gt; правый и левый печёночные протоки -gt; общий печёночный проток —gt; общий жёлчный проток -*¦ двенадцатиперстная кишка.
а. Жёлчные капилляры находятся внутри тяжей гепатоцитов, это тонкие каналы между соседними гепатоцитами. Жёлчные капилляры не имеют собственной стенки, т.к. эти капилляры являются частью межклеточного пространства между соседними гепатоцитами, «запертого» специализированными контактами для предупреждения
Рис. 12-31. Ацинусы печени.
 Выделено два соседних ацинуса. В одном показаны зоны, а в другом — печёночные пластинки. I, 2, 3 — зоны ацинуса, различающиеся по интенсивности кровоснабжения и чувствительности к действию токсинов или к недостатку питательных веществ. В зоне I (центральная часть ацинуса) проходят терминальная ветвь воротной венулы, печёночная артериола и жёлчный проток. Клетки зоны 3 лежат ближе к центральной вене [из Ham А, 1974]
Выделено два соседних ацинуса. В одном показаны зоны, а в другом — печёночные пластинки. I, 2, 3 — зоны ацинуса, различающиеся по интенсивности кровоснабжения и чувствительности к действию токсинов или к недостатку питательных веществ. В зоне I (центральная часть ацинуса) проходят терминальная ветвь воротной венулы, печёночная артериола и жёлчный проток. Клетки зоны 3 лежат ближе к центральной вене [из Ham А, 1974]
просачивания жёлчи в кровь, находящуюся в синусоидах. Жёлчные капилляры слепо начинаются в центральной части классической дольки и идут на её периферию, где впадают в холангиолы.
б. Холангиолы — короткие трубочки на периферии классических долек. Принимают жёлчь из жёлчных капилляров и передают её междольковым жёлчным протокам. Холангиола образована 2-3 холангиоцитами.
- Основные клеточные типы
а. Гепатоциты (рис. 12-32) образуют печёночные пластинки (тяжи). Содержат в изобилии практически все органеллы. Ядро имеет 1-2 ядрышка и, как правило, расположено в центре клетки. 25% гепатоцитов имеет 2 ядра. Для клеток характерна полиплоидия; 55-80% гепатоцитов — тетраплоидны, 5-6% — октаплоидны и только 10% — диплоидны. Хорошо развита гранулярная и гладкая эндоплазматическая сеть. Элементы комплекса Гольджи присутствуют в различных отделах клетки. Количество митохондрий в клетке может достигать 2000. Клетки содержат лизосомы и пероксисомы. Последние имеют вид окружённого мембраной пузырька диаметром до 0,5 мкм. Пероксисомы содержат окислительные ферменты — аминооксидазу, уратоксидазу, каталазу. Как и в митохондриях, в пероксисомах происходит утилизация кислорода. Прямое отношение к образованию этих органелл имеет гладкая эндоплазматическая сеть. Гепатоциты активно накапливают гликоген. В цитоплазме присутствуют многочисленные включения. Маркёры: альбумин, глюкозо-6-фосфатаза, цитокератины 8 и 18, цитохром Р-450, аспартат аминотрансфераза, аланин аминотрансфераза.
б. Эпителий жёлчных протоков (холангиоциты). Маркёры: цитокератины 7 и 19.
в. Эндотелиальные клетки (рис. 12-32) синусоидов имеют удлинённую форму. Пери- нуклеарная область клетки выступает в просвет сосуда. Эндотелиальные клетки контактируют при помощи многочисленных отростков. Между эндотелиальными клетками и гепатоцитами имеется пространство Диссе. В его пределах микроворсинки гепатоцитов соприкасаются с поверхностью эндотелиальных клеток. Ядро расположено вдоль клеточной мембраны со стороны пространства Диссе. В клетке содержатся элементы гранулярной и гладкой эндоплазматической сети. Комплекс Гольджи обычно расположен между ядром и просветом синусоида. В цитоплазме эндотелиальных
Рис. 12-32. Основные клеточные типы печени. Гепатоциты образуют анастомозирующие тяжи. Соприкасающиеся поверхности гепатоцитов формируют жёлчный капилляр. Другой своей поверхностью гепатоциты обращены к синусоиду. Стенка синусоида образована эндотелиальными клетками, между которыми присутствуют клетки фон Купффера, Гепатоциты и эндотелиальные клетки ограничивают пространство Диссе [из Kopf-MaierP, MerkerH-J, 1989]
клеток содержатся многочисленные пиноцитозные пузырьки и лизосомы. Клетки фон Купффера не вступают в контакт с перинуклеарной цитоплазмой эндотелиальных клеток, но располагаются между ними в составе стенки синусоида. Способность эндотелиальных клеток к эндоцитозу значительно менее выражена, чем у клеток фон Купффера. Маркёр эндотелиальных клеток — фактор VIH (фактор фон Вимебранда).
г. Клетки Купффера относятся к системе мононуклеарных фагоцитов. В их цитоплазме содержатся лизосомы, включения железа и пигменты. Характеризуются высокой активностью пероксидазы. Очищают кровь от попавшего в неё чужеродного материала, фибрина, избытка активированных факторов свёртывания крови. Участвуют в фагоцитозе эритроцитов, обмене Hb и жёлчных пигментов. Клетки захватывают железо из крови и аккумулируют его для последующего использования в синтезе Hb. Вместе с гепатоцитами участвуют в инактивации кортикостероидов.
д. Жиронакапливающие клетки (липоциты, клетки Ито) расположены в перисину- соидальном пространстве. Для этих клеток in vitro показана способность синтезировать коллаген, в связи с чем предполагают их участие в развитии цирроза и фиброза печени.
- Функции печени многочисленны.
а. Секреция жёлчи.
б. Синтез белков плазмы (например, альбумины, фибриноген, протромбин, фактор III, липопротеины).
в. Запасание метаболитов (например, гликоген и триглицериды).
г. Глюконеогенез. Превращение аминокислот и липидов в глюкозу.
д. Детоксикация. Инактивация различных лекарственных препаратов и токсических веществ при помощи различных ферментов в ходе реакций окисления, метилирования и связывания.
е. Защита организма
- Фагоцитоз
- Транспорт IgA из пространств Дйссе в жёлчь и далее — в просвет кишки
ж. Кроветворная
- Участие в эмбриональном гемопоэзе [глава 6.1 IV А 2 а]
- Синтез тромбопоэтина
Г. Жёлчный пузырь — растяжимый полый орган грушевидной формы, расположенный под правой долей печени и содержащий 30-50 мл жёлчи. Назначение органа состоит не только в хранении, но и в концентрировании жёлчи за счёт активного транспорта Na* и Cl’ эпителиальными клетками слизистой оболочки.
- Эпителиальные клетки имеют цилиндрическую форму, на апикальной поверхности несут различной величины микроворсинки, покрытые гликопротеиновой оболочкой. Боковая поверхность клеток образует выросты. Среди эпителиальных клеток шейки жёлчного пузыря присутствуют клетки, секретирующие слизь и гормоны.
- Мышечная оболочка представлена гладкомышечными клетками. Холецистокинин, вырабатываемый энтероэндокринными клетками слизистой оболочки тонкой кишки, стимулирует сокращение ГМК и эвакуацию жёлчи. При пустом жёлчном пузыре сокращение мышечной оболочки приводит к образованию складок слизистой.
- Наружная оболочка жёлчного пузыря — серозная. Она покрывает весь орган, за исключением места его прилегания к печени.
Источник